



Héritage ou convergence ? Les chemins sinueux de l’évolution des espèces
PDF
Comment expliquer que certaines espèces se ressemblent plus que d’autres ? Les espèces sont le produit de leur histoire : l’évolution. Elles représentent les feuilles de l’arbre de la vie, qui émerge d’un tronc principal symbolisant leur ancêtre commun à toutes. Deux espèces proches dans cet arbre se sont donc séparées l’une de l’autre il y a fort peu de temps –elles ont un ancêtre commun récent – et souvent se ressemblent. Deux espèces éloignées dans l’arbre sont donc généralement très différentes car elles ont suivi des branches évolutives indépendantes. Certaines paires d’espèces éloignées historiquement mais très ressemblantes morphologiquement constituent toutefois des contre-exemples. En fait, ces ressemblances intrigantes sont le produit de l’adaptation répétée des espèces aux mêmes conditions environnementales. Ces convergences évolutives sont des éléments majeurs permettant d’affirmer que l’adaptation est une force fondamentale de l’évolution du vivant.
Une caractéristique remarquable du monde vivant est la très grande diversité de formes qu’il prend. On regroupe souvent les individus en espèces, ce qui permet de décrire de façon commode cette biodiversité. On observe ainsi sur les clichés de la figure 1 une certaine diversité de formes d’espèces au sein des mammifères. Mais certaines espèces se ressemblent plus que d’autres. Ainsi, l’ours polaire ressemble beaucoup plus à l’ours brun qu’au gorille.
1. Les cousins germains se ressemblent plus que les cousins éloignés
Les espèces ont longtemps été décrites d’un point de vue fixiste, c’est-à-dire comme des entités immuables, des “essences” qui étaient a priori pensées par le divin (lire Qu’est-ce que la biodiversité ?). À la fin du XVIIIe et au début du XIXe siècle, des scientifiques comme Lamarck (1744-1829), Wallace (1823-1913) ou encore Darwin (1809-1882, lire focus Darwin) ont pourtant révolutionné cette vision traditionnelle de la biodiversité en prouvant que les espèces vivantes évoluaient depuis leur origine (Lire Lamarck et Darwin: deux visions divergentes du monde vivant) et qu’elles descendaient donc toutes d’un seul et unique ancêtre commun. En quoi cette découverte nous permet-elle de mieux comprendre les ressemblances entre les espèces ?
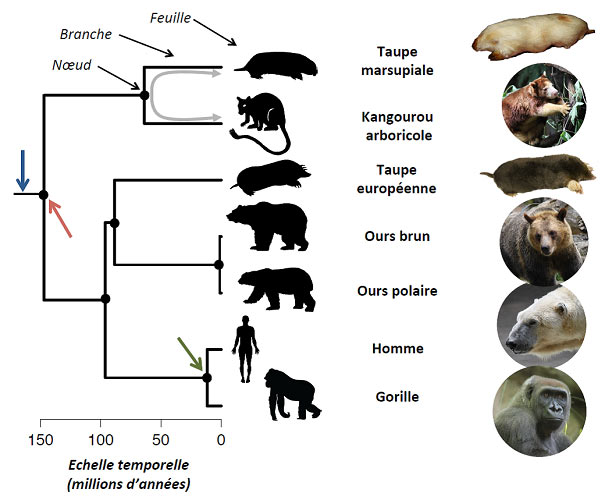
2. Quelques contre-exemples…
Pourtant, il existe des contre-exemples : des espèces éloignées évolutivement peuvent se ressembler de façon frappante, comme c’est le cas entre certains grands groupes d’animaux marins, de plantes ou de mammifères.
2.1. Les animaux marins
Les poissons à nageoires rayonnées (par exemple le thon) [4] et les cétacés (mammifères marins dont font partie les baleines et les dauphins) se sont séparés il y a environ 430 millions d’années [5]. Pourtant, ne partagent-ils pas une allure générale semblable ? À ce sujet, Darwin [6] notait déjà que :
« Des animaux appartenant à deux lignées d’ancêtres très distincts peuvent, en effet, s’être adaptés à des conditions semblables, et avoir ainsi acquis une grande ressemblance extérieure ; mais ces ressemblances, loin de révéler leurs relations de parentés tendent plutôt à les dissimuler. […] Ainsi, la forme du corps et les membres en forme de nageoires […] constituent dans les deux classes une adaptation spéciale en vue d’un mode de locomotion aquatique »
Darwin suggère ici que les deux groupes ont, au cours de l’évolution, développé une forme hydrodynamique et des nageoires ce qui leur permet de se déplacer efficacement dans l’eau.
2.2. Les mammifères souterrains
Des animaux ressemblant à des taupes (Figure 1) existent parmi les mammifères dans des régions différentes du globe. Ainsi, la taupe européenne est présente dans une partie de l’Eurasie alors que la taupe marsupiale est présente en Australie. Ces espèces présentent une forme fouisseuse similaire, adaptée à la vie souterraine (Figure 1). Elles possèdent en effet toutes un corps trapu en forme de “saucisse” -sans cou distinguable- équipé de puissants membres antérieurs dotés de larges griffes permettant de creuser efficacement. Elles ont toutes plus ou moins perdu les organes sensoriels extérieurs (oreille externe, vue). La ressemblance entre ces « taupes » ne provient pas d’un héritage commun, c’est-à-dire d’un ancêtre commun ayant lui-même une forme de taupe. En effet, la taupe marsupiale est phylogénétiquement plus proche d’un kangourou que d’une taupe européenne (Figure 1) qui, elle, est plus proche de l’homme que de la taupe marsupiale. Ainsi, la forme de « taupe » est apparue à plusieurs reprises dans des branches éloignées de l’arbre phylogénétique des mammifères.
2.3. Les plantes en coussin

2.4. Les plantes « cactoïdes »

2.5. Les plantes carnivores

3. Quand la sélection naturelle « brouille les pistes »
Ainsi, bien que certaines espèces puissent être très éloignées phylogénétiquement, elles peuvent avoir convergé vers une morphologie similaire, pour s’adapter à des contraintes environnementales similaires. L’hypothèse d’une évolution aléatoire ayant eu lieu précédemment (hypothèse 1) ne semble donc pouvoir expliquer ces phénomènes d’adaptation du vivant à son environnement. Une contribution majeure de Darwin (1859) a justement été de proposer le mécanisme permettant l’adaptation des espèces : la sélection naturelle.
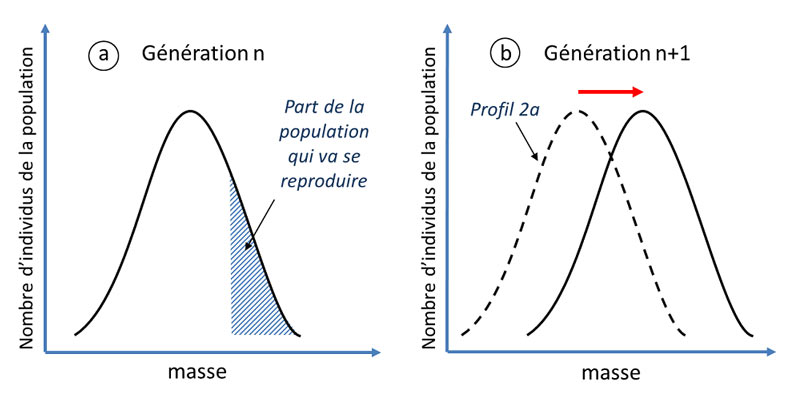
Dans la théorie de Darwin, la sélection naturelle requiert quatre ingrédients :
- La première condition nécessaire est l’existence d’une certaine diversité de formes à l’intérieur même d’une espèce, par exemple des individus de plantes en coussin plus ou moins compacts (figure 5A). C’est justement cette diversité qui était ignorée des fixistes, pour qui elle ne représentait que des « erreurs de copies » de l’essence divine de l’espèce. Cependant, cette diversité est bel et bien réelle : il suffit de regarder la diversité au sein de l’espèce humaine.
- La seconde condition est une reproduction limitée des individus. Imaginez simplement que chaque individu d’une espèce donne deux descendants à chaque génération. Si l’on part d’un seul individu, au bout de vingt générations, la population aura atteint la taille de 220 individus, soit plus d’un million et au bout de cent générations, plusieurs milliards de milliards… ce qui est a priori impossible sur une planète aux ressources importantes, mais limitées. Ainsi, seul un nombre limité d’individus de l’espèce vont contribuer à la génération suivante.
- La troisième condition stipule que le cortège d’individus qui appartiennent au lot qui se reproduit possède des caractéristiques particulières qui leur ont permis de mieux survivre dans un milieu aux contraintes données et donc de davantage se reproduire. Par exemple, dans un environnement de plus en plus rude, on peut imaginer que seuls les individus de plantes en coussins les plus compacts survivent assez longtemps pour se reproduire (Figure 5A).
- Si ces caractéristiques sont héritables, alors elles vont petit à petit se répandre dans la population et l’espèce va progressivement évoluer, au fil des générations (Figure 5B).
Darwin fournit ici une théorie permettant d’expliquer l’adaptation des espèces à leur environnement. Ainsi, si deux branches éloignées de l’arbre subissent les mêmes contraintes de l’environnement –comme un déficit en azote ou un climat rude- elles pourront conduire à des espèces morphologiquement proches, mais phylogénétiquement éloignées : plantes carnivores et plantes en coussins, respectivement. En définitive, les espèces sont des mosaïques de caractères qui ont pu prendre des chemins évolutifs différents. Par exemple, toutes les plantes en coussins présentent la même forme générale, mais possèdent des morphologies florales bien distinctes, caractéristiques de leur famille (Figure 3). Les cétacés possèdent des poumons -hérités d’un lointain ancêtre vertébré- emballés dans une morphologie hydrodynamique qui elle n’est pas héritée, mais bien convergente.
4. Synthèse
Les ressemblances entre espèces peuvent donc être le fruit d’un héritage historique ou d’une convergence évolutive. Pour être capable de distinguer entre les deux hypothèses, il faut donc disposer d’une phylogénie des espèces et de mesure de leurs caractéristiques morphologiques, anatomiques ou physiologiques. Historiquement, les arbres phylogénétiques étaient construits à partir des données morphologiques ou anatomiques, ce qui pouvait poser problème si l’on se servait de caractères convergents pour les construire. Dans ce cas, on risquait en effet de regrouper des espèces qui ont en fait convergé et dont les ressemblances ne sont pas héritées. Les dernières décennies ont vu l’avènement du séquençage de la molécule d’ADN et de son utilisation en phylogénie. Les ressemblances moléculaires sont beaucoup moins sujettes à ces convergences évolutives, et si elles sont correctement utilisées elles sont donc un outil de choix pour reconstruire les relations de parenté entre espèces. Ainsi, c’est bien la comparaison entre les classifications taxonomiques plus classiques basées sur la morphologie et les classifications phylogénétiques issues des données moléculaires qui vont permettre de mettre en lumière les convergences évolutives. C’est, en quelque sorte, une synergie entre les anciennes et les nouvelles approches en évolution.
Références et notes
Photo de couverture : Photo de groupe de dauphins prise à Xcaret, Mexique © Truncatus [domaine public] via Wikimedia Commons
[1] Darlu P. & Tassy P. (1993) Reconstruction phylogénétique. Concepts et Méthodes. Coll. “Biologie théorique”, no 7. Masson, Paris. (Livre en ligne sur le site de la Société Française de Systématique : http://sfs.snv.jussieu.fr/index.php/la-reconstruction-phylogenetique-concepts-et-methodes/).
[2] Bininda-Emonds O.R., Cardillo M., Jones K.E., MacPhee R.D., Beck R.M., Grenyer R., Price S.A., Vos R.A., Gittleman J.L. & Purvis A. (2007) The delayed rise of present-day mammals. Nature, 446, 507-512.
[3] Les dates mentionnées dans cet article proviennent de l’étude de Bininda-Edmonds et al. (référence 2). Ces dates sont spéculatives et reflètent l’état des connaissances actuelles en phylogénie des mammifères, elles sont donc amenées à être modifiée au gré des nouvelles découvertes.
[4] Les « poissons » regroupent les poissons cartilagineux (requins et raies), les poissons à nageoires rayonnées (par exemple la truite ou le thon), les myxines, les lamproies et les dipneustes. Ceux-ci ne forment plus un groupe utilisé en systématique (mais –à juste titre- encore en cuisine), car l’ancêtre commun des « poissons » ne leur est pas exclusif. Par exemple, l’ancêtre commun à la truite et au requin est aussi l’ancêtre commun de l’homme. Les « poissons » ne représentent donc pas une lignée évolutive propre, mais bien une concaténation de plusieurs groupes distincts.
[5] http://www.timetree.org. Ces dates sont spéculatives et reflètent l’état des connaissances actuelles en phylogénie, elles sont donc amenées à être modifiée au gré des nouvelles découvertes.
[6] Darwin C. (1859) On the origins of species by means of natural selection. London: Murray, 247.
[7] voir aussi www.cushionplants.eu
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Pour citer cet article : MAZEL Florent, LAVERGNE Sébastien, DOUZET Rolland, THUILLER Wilfried (2 février 2019), Héritage ou convergence ? Les chemins sinueux de l’évolution des espèces, Encyclopédie de l’Environnement. Consulté le 11 avril 2025 [en ligne ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/vivant/heritage-convergence-chemins-sinueux-de-levolution-especes/.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.


