



Pourquoi le moustique tigre est-il aussi invasif ?
PDF
D’où proviennent les espèces invasives ? Comment sont-elles introduites ? L’intensification des échanges commerciaux a conduit à une explosion de la fréquence d’introduction d’insectes exotiques à l’échelle globale. De nombreux facteurs sont néanmoins déterminants pour le succès d’une invasion biologique, en particulier les caractéristiques écologiques et génétiques des populations introduites. Ainsi, la plupart des introductions d’espèces se soldent par un échec. La mondialisation augmente la probabilité d’introduire une espèce prédisposée à l’invasion, au bon moment, et au bon endroit. L’analyse génétique des populations invasives donne de précieuses informations sur les modalités d’introduction et d’expansion, à l’échelle globale et à l’échelle très locale du paysage.
- 1. Biologie de l’espèce
- 2. D’où viennent les populations invasives ?
- 3. Une expansion rapide en Europe
- 4. Une espèce prédisposée au transport et aux climats tempérés
- 5. Une dissémination passive via le transport routier
- 6. Une expansion naturelle rapide grâce à l’urbanisation des territoires
- 7. Messages à retenir
1. Biologie de l’espèce
Une espèce invasive est une espèce dont certaines populations ont été transportées à partir d’une région donnée vers une autre région géographiquement distante (populations introduites), et qui une fois introduites sont capables de se reproduire (populations établies) et d’étendre leur distribution géographique (populations invasives). Les mécanismes à l’œuvre durant ce processus d’invasion sont indépendants de la notion d’impacts, qu’ils soient d’ordre écologique, sanitaire, ou économique. Néanmoins, les invasions biologiques sont reconnues comme l’une des principales causes des changements de la biodiversité [1].
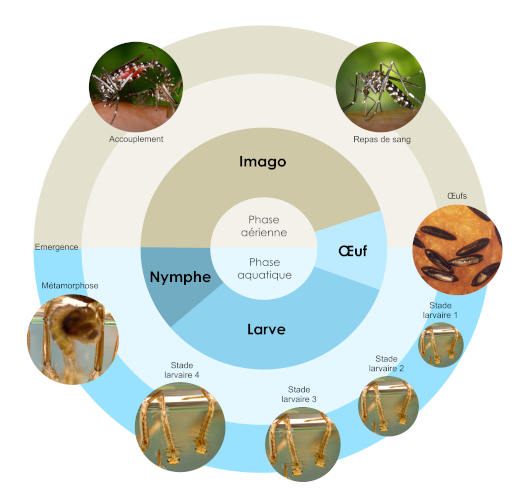
1.1. Cycle de vie
Le cycle de vie du moustique tigre comprend quatre stades : œuf, larve, nymphe, et imago (adulte), répartis sur deux phases en fonction du milieu dans lequel les stades se produisent (Figure 1). La première phase, aquatique, comprend les stades œuf, larve, et nymphe, le stade adulte étant aérien [4]. Les moustiques adultes mâle et femelle se nourrissent de nectar, mais les femelles ont besoin d’un repas de sang pour apporter les nutriments nécessaires au développement des œufs. Les œufs sont pondus sur une surface solide proche de la surface de l’eau, et poursuivent leur développement une fois que le support est recouvert d’eau. Les œufs sont résistants à la sécheresse.
1.2. Écologie

- des récipients naturels, tels que les souches d’arbres, ou les Broméliacées ;
- des réservoirs artificiels, tels que les pneus usagés, les récipients abandonnés, soucoupes de pots de fleurs, ou récupérateurs d’eaux pluviales.
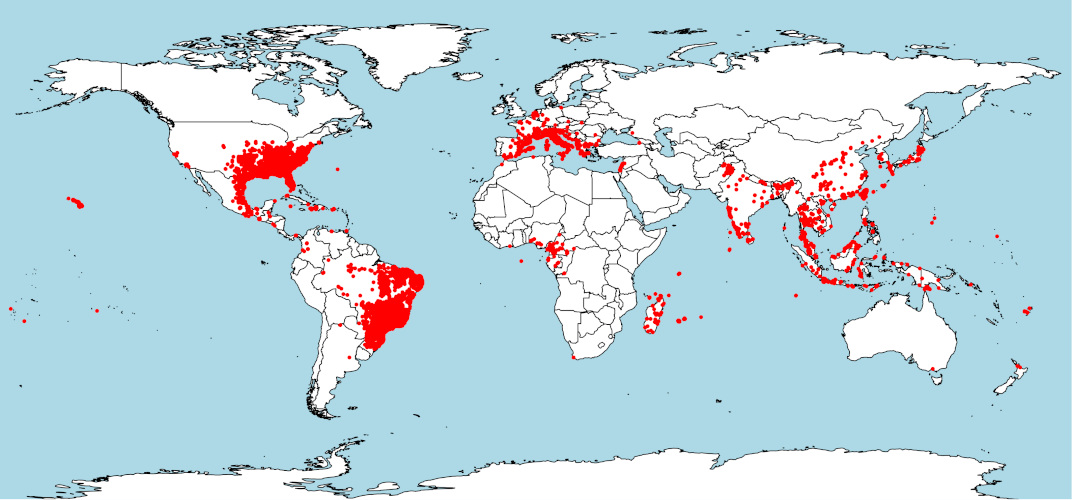
Dans son aire d’origine en Asie, le moustique tigre est présent dans des environnements très contrastés : des régions tropicales d’Asie du Sud-Est (larges îles Indonésiennes, Péninsule Malaise) aux régions tempérées du Nord-Est de la Chine et du Japon, en passant par les régions subtropicales de la Péninsule Indochinoise jusqu’à l’Inde plus à l’Ouest (Figure 3). [5]
2. D’où viennent les populations invasives ?
Le taux d’introduction d’espèces exotiques a considérablement augmenté durant les dernières décennies [2], notamment à cause de l’accélération des échanges commerciaux et du transport des marchandises et des personnes (Figure 4).
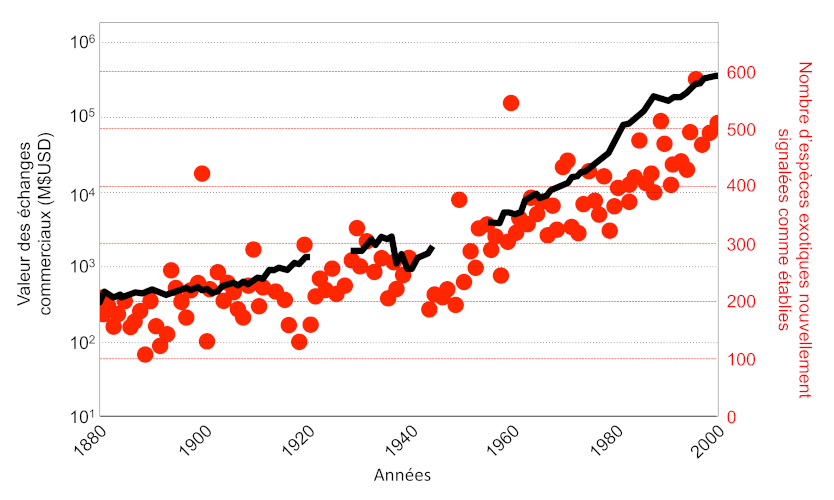
La croissance rapide du commerce de marchandises à l’échelle mondiale a multiplié les introductions du moustique tigre. Sa diffusion a probablement débuté à la fin du 19e siècle, mais sa répartition s’est rapidement étendue à l’ensemble des continents excepté l’Antarctique au cours des dernières décennies, notamment en Europe où l’espèce est présente depuis la fin des années 1970 [9].
L’analyse génétique des populations européennes montre trois introductions indépendantes : en Albanie à partir de la Chine, au Nord de l’Italie à partir des États-Unis, et au Centre de l’Italie, où la variabilité génétique des populations résulte d’un mélange entre celles initialement présentes au Nord de l’Italie et une nouvelle introduction en provenance de la Chine [10]. Les patrons de migration à l’échelle globale démontrent que les populations sources à l’origine des introductions correspondent aux partenaires commerciaux internationaux au moment des premières introductions.
Bien que chaque invasion biologique ait sa propre histoire, les introductions d’autres espèces d’insectes, telles que la coccinelle [11] et le moucheron [12] asiatiques, se sont déroulées selon un schéma géographique similaire. L’introduction en Europe s’est faite depuis l’Asie via des populations tête de pont aux États-Unis, suggérant un rôle prépondérant des réseaux de transport par l’Homme.
3. Une expansion rapide en Europe
3.1. Origine des populations
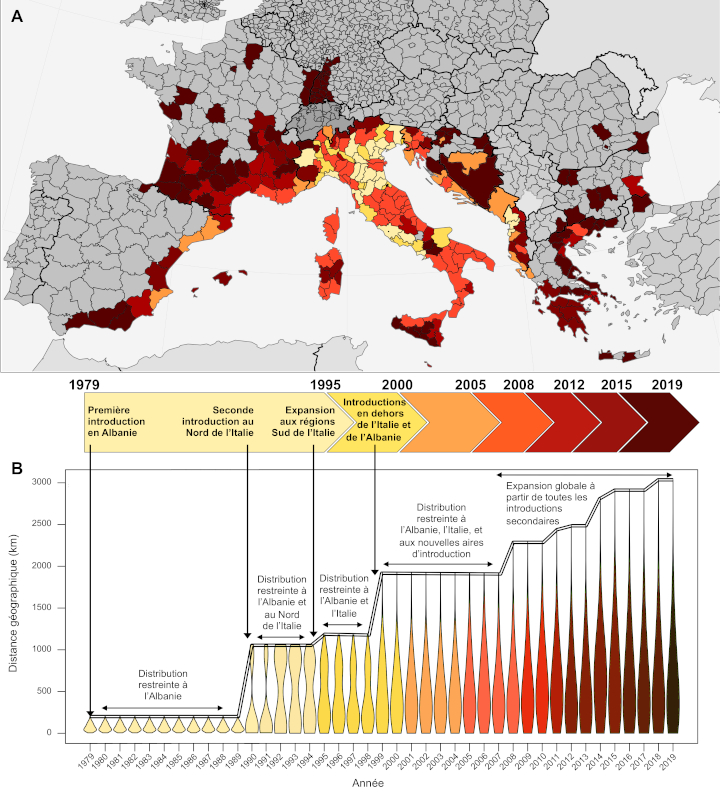
L’analyse d’un échantillonnage représentatif de la distribution de l’espèce en Europe a permis de reconstruire l’histoire précise de l’expansion. Les trois populations initialement introduites en Europe (Albanie, Nord de l’Italie, Centre de l’Italie) ont constitué des centres de dispersion (Figure 6). Les voies de migration reflètent la géographie des transports humains : les échanges génétiques corrèlent au contexte géopolitique dans le bassin Méditerranéen.
3.2. Nombre d’introductions
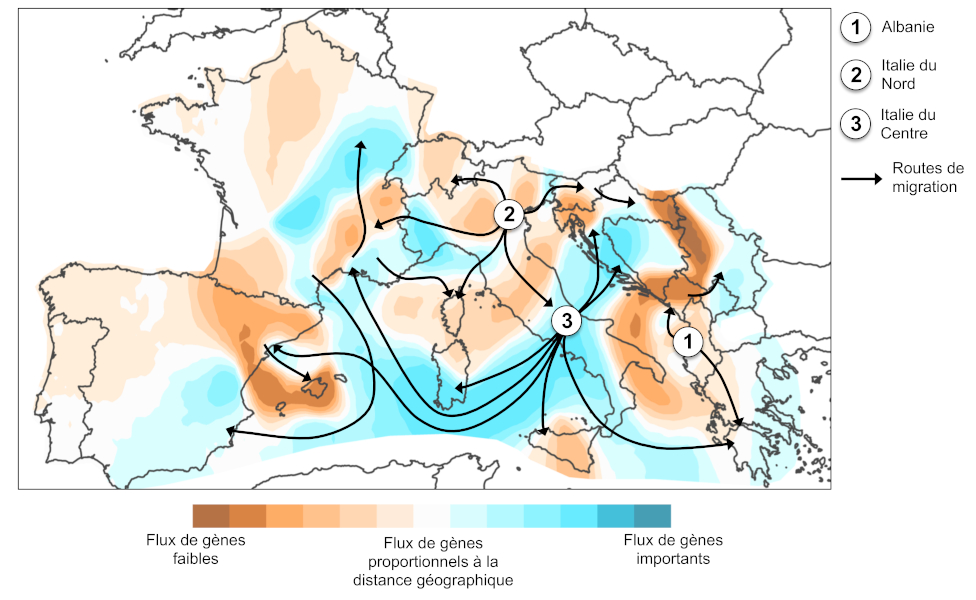
Les introductions multiples constituent un des principaux facteurs augmentant la diversité génétique dans les populations [15]. Elles permettent la mise en contact de deux groupes d’individus génétiquement distincts, dont le brassage conduit à l’apparition de nouvelles combinaisons génétiques. Ainsi, les populations du moustique tigre établies au Centre de l’Italie, issues du brassage entre deux populations sources, présentent des niveaux de diversité génétique plus élevés que ceux enregistrés dans l’aire native de l’espèce [10]. Par ailleurs, la forte connectivité entre les populations introduites a probablement favorisé le maintien d’une forte diversité génétique au sein des populations sur les fronts d’expansion, contribuant à l’expansion rapide à l’échelle de l’Europe (Figure 5).
4. Une espèce prédisposée au transport et aux climats tempérés
Les échanges commerciaux déterminent l’origine des populations introduites et le nombre d’introductions. Ils ne peuvent en revanche à eux-seuls expliquer le succès du moustique tigre. Différents éléments de sa biologie et de son écologie ont également joué un rôle dans les succès d’introduction et d’établissement.
4.1. Probabilité d’introduction
L’adaptation des organismes aux habitats modifiés par l’homme peut accroître leur probabilité d’introduction dans une nouvelle région car les populations situées dans des zones fréquentées par l’homme sont plus susceptibles d’être transportées [16]. Parmi les gîtes larvaires qu’utilise le moustique tigre, les pneus usagés constituent le site de ponte préférentiel. De par leur résistance à la sécheresse, les œufs peuvent survivre durant plusieurs mois et être transportés sur de longues distances. L’intensification du commerce des pneus usagés a donc initié l’expansion mondiale de l’espèce via le transport des œufs.
4.2. Probabilité d’établissement
La distribution du moustique tigre en régions tempérées correspond à certains seuils de conditions climatiques [17] :
- une température moyenne du mois le plus froid supérieure à 0°C pour la survie hivernale des œufs ;
- une température moyenne annuelle de 11°C pour le maintien de l’activité des adultes ;
- un minimum de 500 mm de précipitations annuelles pour le maintien en eau des sites de ponte.
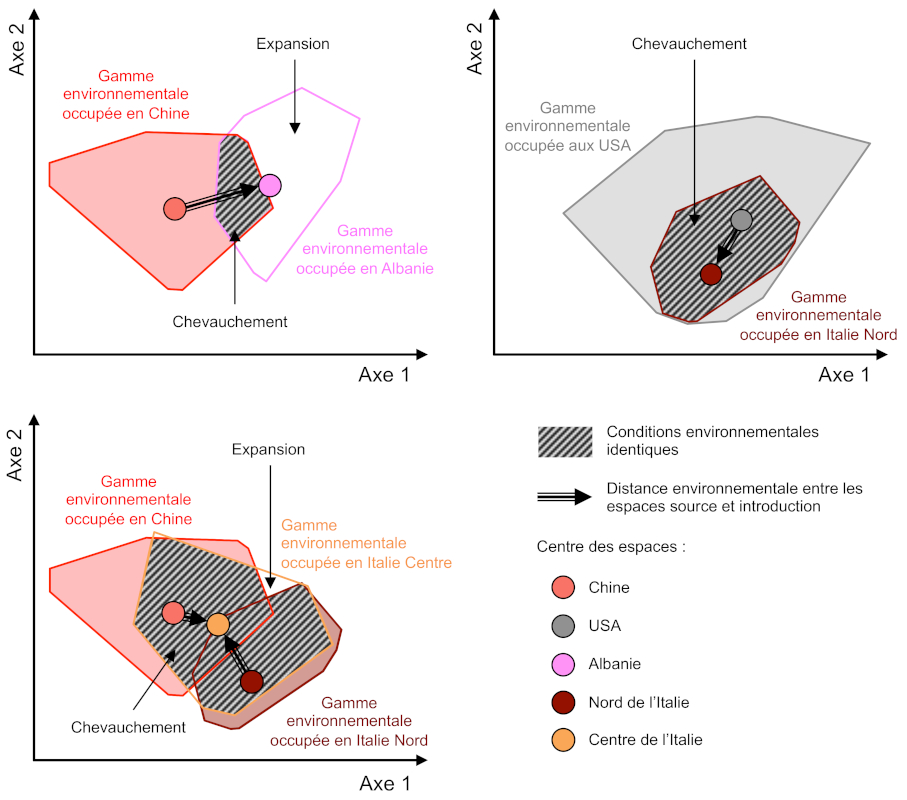
Comme chez de nombreux insectes, la survie hivernale du moustique tigre dans des environnements froids est principalement déterminée par la capacité des œufs à entrer en diapause. Cette adaptation physiologique est déjà présente dans les régions tempérées d’Asie qui sont à l’origine de l’introduction en Albanie et au Centre de l’Italie, ainsi qu’aux États-Unis, à l’origine de l’introduction au Nord de l’Italie [5],[18],[19]. Ainsi, le moustique tigre était déjà pré-adapté pour s’installer en Europe. De plus, les contraintes environnementales rencontrées lors de l’introduction en Europe étaient peu différentes de celles rencontrées dans les populations sources [20], pouvant expliquer l’établissement rapide des populations (Figure 7). Ces contraintes environnementales incluent les contraintes climatiques (température, précipitations) mais également anthropiques (urbanisation et artificialisation des milieux).
5. Une dissémination passive via le transport routier
Il existe une relation de cause à effet directe entre la dispersion [23] et les flux de gènes entre populations. Dès lors, la génétique du paysage [24] cherche à identifier des facteurs géographiques et paysagers favorisant la connectivité des populations. Contrairement à la reconstruction des routes de colonisation qui s’étudie à une échelle globale, la connectivité des populations doit être appréhendée à une échelle locale. Bien que la dispersion à longue distance assistée par l’homme puisse être caractérisée par des « sauts », les ilots colonisés sur les fronts d’expansion offrent d’excellents laboratoires naturels pour étudier les facteurs paysagers impactant la connectivité des populations.
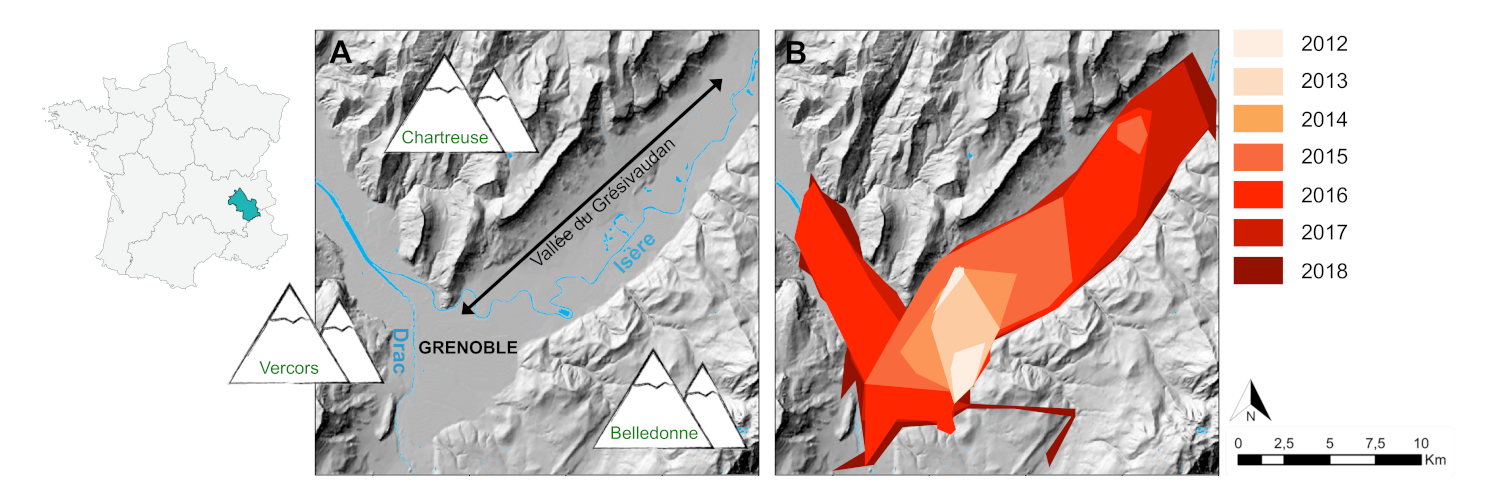
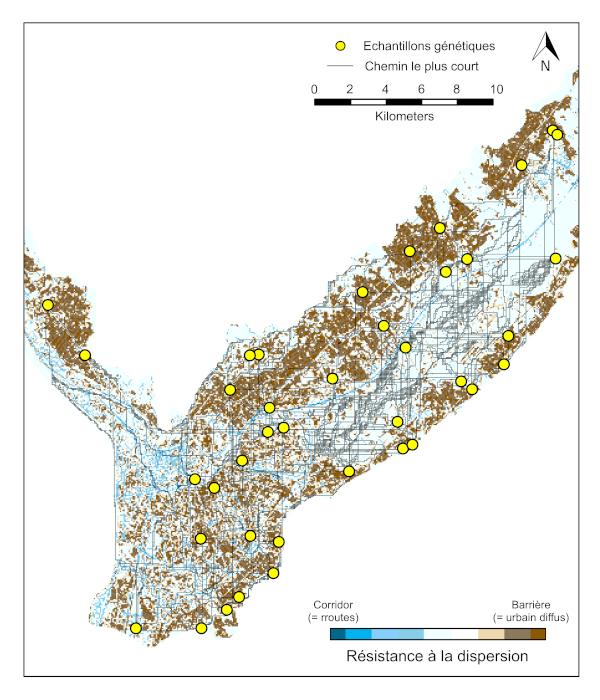
La modélisation des facteurs structurant la variabilité génétique des populations à l’échelle du paysage Grenoblois, utilisant différents types d’habitats (milieux ouverts, forêts, aires urbaines denses, aires urbaines résidentielles, cours d’eau, réseaux routiers), révèle que la dispersion passive du moustique tigre le long des axes routiers induit une forte connectivité entre des populations géographiquement distantes de part et d’autre de Grenoble [25] (Figure 9).
6. Une expansion naturelle rapide grâce à l’urbanisation des territoires
Dans la région de Grenoble, la présence du moustique tigre est déterminée par (Figure 10):
- Une température moyenne annuelle minimale de 11°C, corrélée à une température optimale durant les mois estivaux, période d’activité des adultes, et à une température hivernale permettant la survie des œufs. Ce seuil de température correspond à l’isocline 800 m.
- La présence d’habitats urbains diffus, correspondant aux zones résidentielles avec des jardins où les sites de pontes (coupelles de pots de fleurs, récupérateurs d’eau) et les humains (repas de sang pour les femelles) sont en quantité abondante permettant d’accomplir le cycle de reproduction.

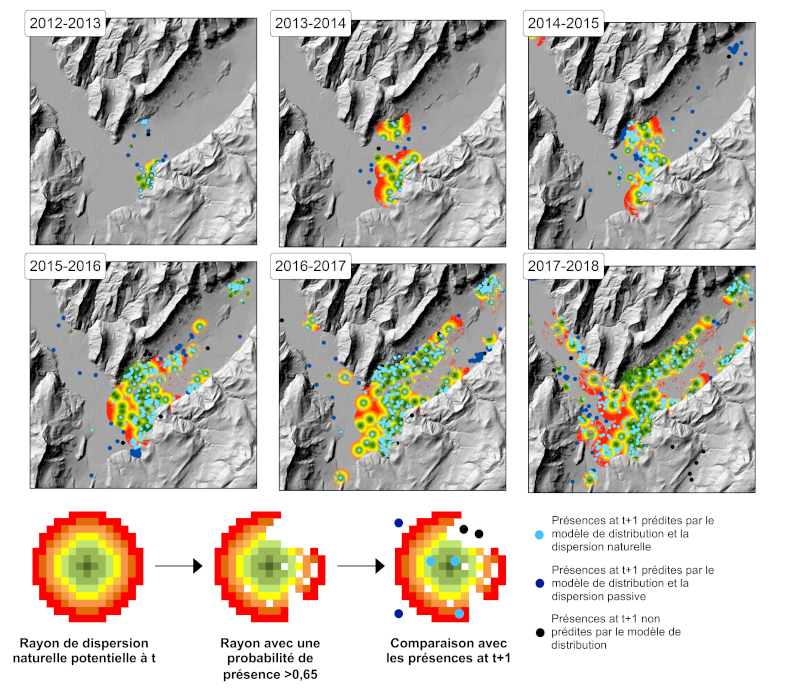
La proportion importante d’habitats urbains diffus continus et favorables pour le moustique tigre dans la région de Grenoble a favorisé la colonisation rapide de la quasi-totalité des zones résidentielles quelques années après son introduction. La croissance démographique humaine et l’attrait pour les zones résidentielles sur les pourtours des zones urbaines denses induisent progressivement une extension des espaces résidentiels urbains et un remplacement des milieux ouverts, tels que les prairies ou les terrains agricoles, offrant de nouveaux habitats favorables à l’établissement et donc l’expansion du moustique tigre. Les changements d’utilisation des terres et les pressions exercées par l’homme sur les écosystèmes favorisent l’établissement et accélèrent l’expansion des espèces invasives (lire aussi : Quand les plantes envahissantes se promènent aussi dans les champs).
7. Messages à retenir
- L’analyse génétique des populations invasives permet de retracer leur histoire (introduction, établissement, expansion) à l’échelle globale, mais également à l’échelle très locale du paysage.
- Le nombre d’introductions et l’origine des populations introduites déterminant leur capacité d’installation (diversité génétique et traits adaptatifs) sont conditionnés par les réseaux de transport par l’Homme.
- La dispersion longue distance assistée par l’homme le long des axes autoroutiers induisant des échanges fréquents entre les populations favorise leur persistance à long terme.
- Le rôle de la dispersion naturelle à l’échelle du paysage dans l’expansion spatiale des populations est souvent sous-estimé et dépend de la quantité d’habitats favorables.
- L’étendue et la disponibilité de ces habitats sont directement influencées par le changement d’utilisation des terres.
Notes et références
Image de couverture. Moustique-tigre. [Source : James Gathany, CDC / Public domain]
[1] Simberloff, D. et al. (2013). Impacts of biological invasions: what’s what and the way forward. Trends in Ecology and Evolution, 28(1), 58−66.
[2] Seebens, H. et al. (2017). No saturation in the accumulation of alien species worldwide. Nature Communications, 8(1), 1−9.
[3] S. Lowe, M. et al. (2007). 100 espèces exotiques envahissantes parmi les plus néfastes au monde. Une sélection de la Global Invasive Species Database. Groupe de spécialistes des espèces envahissantes (ISSG).
[4] Syndicat de Lutte contre les Moustiques du Bas-Rhin. Moustique tigre – Biologie.
[5] Sherpa, S. (2019). Histoire de la colonisation et déterminants du succès invasif des populations du moustique tigre en Europe. Thèse de doctorat, Université Grenoble Alpes.
[6] La diapause est un arrêt temporaire du développement, facultatif ou obligatoire, et généralement déterminé par les conditions environnementales (température et photopériode).
[7] Estoup, A., & Guillemaud, T. 2010. Reconstructing routes of invasion using genetic data: why, how and so what? Molecular Ecology, 19(19), 4113−4130.
[8] Estoup, A. & Guillemaud, T. (1999). Retracer l’invasion pour mieux la combattre.
[9] European Center for Disease Prevention and Contrôle (ECD). Aedes albopictus.
[10] Sherpa, S. et al. (2019). Unraveling the invasion history of the Asian tiger mosquito in Europe. Molecular Ecology, 28(9), 2360−2377.
[11] Institut National de la Recherche Agronomique (INRA) (2010). Les routes d’invasion de la coccinelle asiatique Harmonia axyridis retracées.
[12] Fraimout, A. et al. (2017). Deciphering the routes of invasion of Drosophila suzukii by means of ABC random forest. Molecular Biology and Evolution, 34(4), 980−996.
[13] Bock, D. G. et al. (2015). What we still don’t know about invasion genetics. Molecular Ecology, 24(9), 2277−2297.
[14] Simberloff, D. 2009. The role of propagule pressure in biological invasions. Annual Review of Ecology, Evolution, and Systematics, 40, 81−102.
[15] Dlugosch, K. M. & Parker, I. M. (2008). Founding events in species invasions: genetic variation, adaptive evolution, and the role of multiple introductions. Molecular Ecology, 17(1), 431−449.
[16] Hufbauer, R. A. et al. (2012). Anthropogenically induced adaptation to invade (AIAI): contemporary adaptation to human‐altered habitats within the native range can promote invasions. Evolutionary Applications, 5(1), 89−101.
[17] Fischer, D. (2014). Climatic suitability of Aedes albopictus in Europe referring to climate change projections: comparison of mechanistic and correlative niche modelling approaches. Eurosurveillance, 19, 20696.
[18] Armbruster, P. A. (2016). Photoperiodic diapause and the establishment of Aedes albopictus (Diptera: Culicidae) in North America. Journal of Medical Entomology, 53(5), 1013−1023.
[19] Sherpa, S. et al. (2019). Cold adaptation in the Asian tiger mosquito’s native range precedes its invasion success in temperate regions. Evolution, 73(9), 1793−1808.
[20] Sherpa, S. et al. (2019). Predicting the success of an invader: Niche shift versus niche conservatism. Ecology and Evolution, 9(22), 12658−12675.
[21] Scholte, E. J. & Schaffner, F. (2007). Waiting for the tiger: establishment and spread of the Aedes albopictus mosquito in Europe. In Emerging pests and vector-borne diseases in Europe, Eds. Takken W., & Knols, B. G. J., Wageningen Academic Publishers. pp. 241−260.
[22] Eritja, R. et al. (2017). Direct evidence of adult Aedes albopictus dispersal by car. Scientific Reports, 7(1), 1−15.
[23] Distance moyenne parcourue entre la naissance et la reproduction d’un individu. La dispersion naturelle dépend de la capacité de déplacement des individus d’une espèce dans un environnement donné, alors que la dispersion passive implique l’intervention humaine dans le déplacement des individus.
[24] Manel, S. et al. (2003). Landscape genetics: combining landscape ecology and population genetics. Trends in Ecology and Evolution, 18(4), 189−197.
[25] Sherpa, S. et al. (2020) Landscape does matter: disentangling founder effects from natural and human‐aided post‐introduction dispersal during an ongoing biological invasion. Journal of Animal Ecology, 89(9), 2027–2042..
[26] Marini, F. et al. (2019). Estimating spatio-temporal dynamics of Aedes albopictus dispersal to guide control interventions in case of exotic arboviruses in temperate regions. Scientific Reports, 9(1), 1−9.
[27] Temps moyen nécessaire pour que les individus d’une génération produisent une nouvelle génération d’individus.
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Pour citer cet article : SHERPA Stéphanie (9 mars 2021), Pourquoi le moustique tigre est-il aussi invasif ?, Encyclopédie de l’Environnement. Consulté le 3 avril 2025 [en ligne ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/vivant/pourquoi-moustique-tigre-invasif/.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.


