





恢复热带稀树草原和热带草地生态系统

当人们想到热带稀树草原时,脑海里会出现大象、牛羚、长颈鹿或狮子在广阔非洲大地上漫游的景象。高草草原随着季节变化从绿色到黄色转变,草丛中或密或稀地散布着乔木和灌木。这类草地生态系统占据了地球表面20%的土地,分布在地球的热带地区,除了非洲,还有美洲和亚洲。由于经常发生大火,以及食草动物的存在(其数量也受到捕食者的控制),限制了乔木和灌木的扩张,从而维持了环境的开放性。由于热带稀树草原植被状况极为复杂,气候变化和土地利用对其影响具有高度不确定性。热带稀树草原已经在很大程度上受到人类活动的影响:如农业用地开垦、城市化占用等,在碳补偿的框架下实施的植树工程也影响了热带稀树草原。热带稀树草原因人类活动而退化,其自然恢复力很低,因而必须实施恢复措施,而且这将是一个持久的挑战。
1. 热带原生草地生态系统
就像原生热带森林和赤道森林(又称为原始森林)一样,原生或原始热带草地生态系统是经过几个世纪才形成的物种极其丰富的生态系统类型[1]。这些原始或古老的草地生态系统包括稀树草原和热带草地,它们通常被统称为“萨王纳草原”(savannas),本文在后面的部分也统一使用“萨王纳”指代。

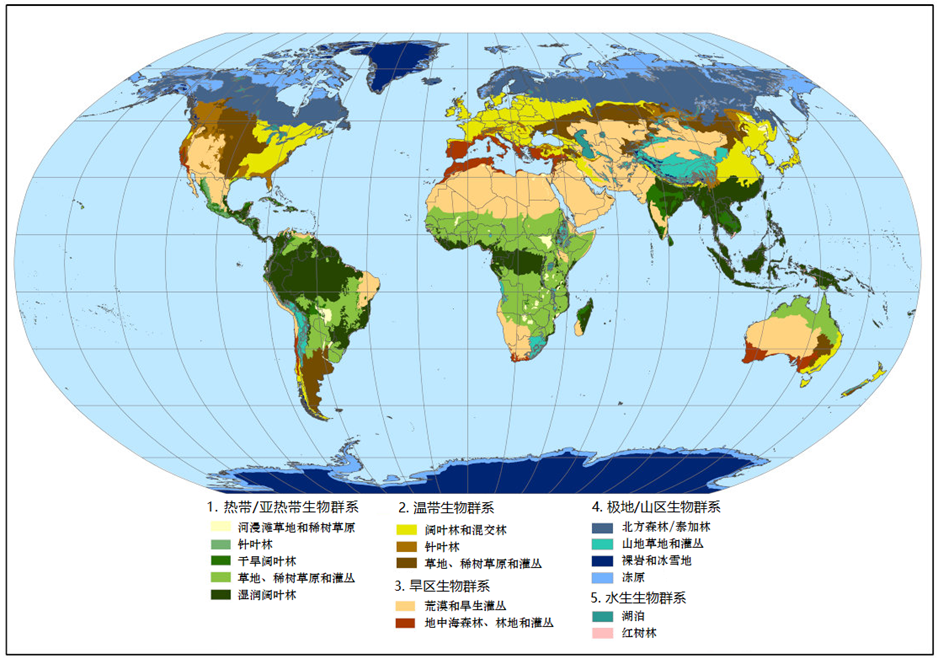
萨王纳草地的类型多样,面积约占陆地表面的20%,在整个热带地区均有分布(图1)。一提到萨王纳这个词,就会让人联想非洲,确实如此,它占据了整个非洲大陆面积的33.5%,养育了那里极具象征性的巨型动物群,包括大象、狮子、斑马 、野牛、长颈鹿和犀牛。全球的萨王纳分布如下(图1~4):
- 非洲的萨王纳主要分布于撒哈拉以南地区、东非地区、中非,甚至远至南非(图2);
- 南美洲和中美洲(图3)萨王纳主要分布于大陆的中部;巴西的萨王纳分布地区被当地人称为“塞拉多”(Cerrado),委内瑞拉和哥伦比亚的萨王纳则被称为“亚诺斯”(Llanos)。在亚马孙雨林的中部和圭亚那也有分布;
- 澳大利亚的萨王纳主要分布在北部和新几内亚南部;
- 在亚洲,萨王纳主要分布于印度和中国,但是面积小,不太为人所知。

这里我们谈论的是热带地区的生态系统,但其他草地生态系统也与萨王纳有着相似的特征,其中一点就是火灾频繁发生,比如位于亚热带的巴西南部、乌拉圭和阿根廷的潘帕斯草原,以及北美的橡树稀树草原(橡树萨王纳)。
2. 萨王纳的结构
萨王纳的地貌多种多样,有些非常开阔,灌木稀少或不存在;而另一些恰恰相反,数目种类多,形成了相对封闭的树冠层(图3)。这些生态系统的共同特征是存在一个主要由C4*草本植物(详见光合作用的碳代谢途径)和非木质草本植物(又称杂类草)连续的草本层。
2.1. 萨王纳生态系统结构的起源
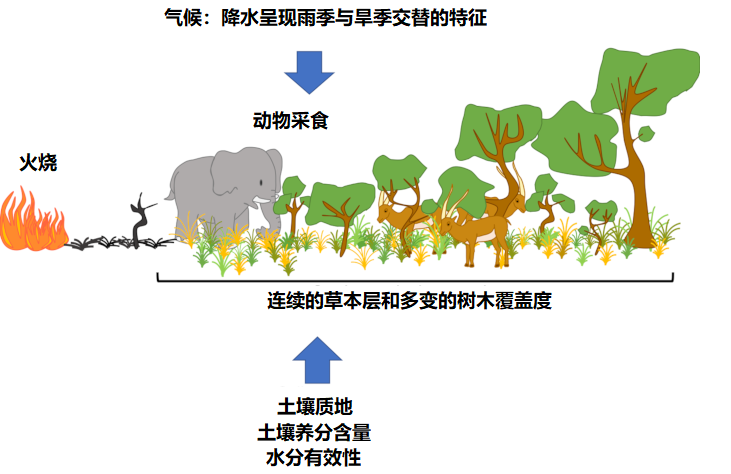
树木通常能抵御火烧,或者在火灾发生后有一定恢复力,但是草本层中的树种往往不耐荫[2]。因此,树木和草本植物在萨王纳中共存就显得不同寻常,这是与水资源、土壤性质和干扰机制(如火灾)诸多过程相互作用的结果,也是这些生态系统植被结构的起源[2],[3]:
- 气候条件(尤其是降雨量和雨季的时长),因为树木生长必须满足其最小降雨量需求[4],[5],[6]。热带地区气候的特点是干湿季节分明[4],[6],许多热带地区分布着大量的萨王纳——开阔的生态系统,单凭降雨量判断,这类地区应该有大片森林[2],[5],[7]。
- 长期干扰,包括火灾和食草动物采食抑制了树木盖度,使萨王纳结构得以维持(图5) 。
- 土壤性质,比如土壤成分和质地会影响植被类型:在持水性能较弱的沙质土壤可以生长较多的灌木,而在持水能力较强的黏质土壤上更多生长的是草本植物(见图2)。

萨王纳等热带草地生态系统是一种古老的生态系统,在火灾和大型食草动物共存的情况下经历了数百万年的演变。C4草本植物起源于渐新世末期,到中新世逐渐占据了主导地位,证明这类生态系统有久远的历史,其物种丰富度和高比例的特有种也是有力的证据。一些热带草地生态系统还生长着一些见证了漫长进化历程的特殊物种[8]。
2.2. 火——塑造萨王纳生态系统的主要力量
仅凭气候特征不能准确推断萨王纳的分布。食草动物(特别是“巨型食草动物”,如大象和有蹄类动物)和火灾干扰在萨王纳的生态和进化中发挥着主要作用[2],[5],[9](见图2~5)。
萨王纳生态系统草本层中的草类作为一种主要生物燃料促进了火灾的发生,同时又作为遮蔽物抑制了乔木和灌木的生长。草类和一般草本物种的生命周期适应了火灾,能够在火灾后非常迅速地再生和繁殖(图6),因此,它们在萨王纳生态系统中的存在有赖于火灾反复出现。

- 萨王纳的树木表现出一些耐火的相关特征,特别是厚的树皮,如在塞拉多和亚诺斯生长的美洲锡叶树(Curatella americana,五桠果科)、巴西油桃木(Caryocar brasiliense,油桃木科)、风铃木属乔木(Handroanthus ochraceus,紫葳科)、翼齿豆属乔木(Pterodon emarginatus,豆科)、鲍迪豆属乔木(Bowdichia virgilioides,豆科)和番荔枝属乔木(Annona crassiflora,番荔枝科);
- 相比而言,森林乔木的树皮要薄得多,易受火烧伤害,如柯柏胶树(Copaifera langsdorffii,豆科)、铁木豆属乔木(Swartzia flaemingii,豆科)、孪叶豆(Hymenaea coubaril,豆科)、翅玉蕊属乔木(Cariniana estrellensis,玉蕊科)、芳香木瓣树属乔木(Xylopia aromatica,番荔枝科)。
在热带地区,若没有火灾等因素的干扰,许多萨王纳的降雨量足以形成森林[11]。在这些地区,森林和萨王纳是生物群落的两种状态(state)[5]、[7],其环境条件既可能出现森林,也可能出现萨王纳,实际出现哪一种植被主要取决于干扰因素,特别是干扰的强度和频率。
3. 生物多样性和保护问题
萨王纳是一种特殊的遗产,有着极其丰富的生物多样性,特别是有丰富的地方特有物种。例如,被称为塞拉多的巴西萨王纳,其面积超过200万平方公里,有超过12 000种植物,是世界上物种最丰富的萨王纳(见《焦点:塞拉多生物群系》)。
非洲的萨王纳除了具有多样的植被类型外,还是代表性巨型动物的家园,大象、斑马、长颈鹿、狮子和猎豹的家园。此外,萨王纳也是人类文化遗产和历史的一部分。例如,一些研究表明人类天生偏爱开阔的景观,这可能源于人类在东非萨王纳的长期演化过程,这些环境通常被认为是人类进化的驱动力之一。
然而,人类活动即使不是影响萨王纳的起源,但随着时间的推移也深刻地改变了火灾和动物的采食模式,进而影响了萨王纳的分布[12]。目前许多人仍然生活在这些生态系统的中心,并依赖其提供的生态系统服务,如清洁水质、提供放牧场所和狩猎机会等。
热带草地生态系统是文化和自然遗产,但是当前它们受到严峻的威胁,包括大面积土地转为农用地、控制火灾的政策以及为商业目的或为碳补偿进行的植树造林。例如,亚马孙地区塞拉多草地转变为农业用地的比例比该地区森林转化为农业用地的比例要高得多[13],[14]。
4. 萨王纳的恢复力

萨王纳对自然干扰有极强的恢复力,甚至依赖这些干扰维持其开阔的地貌特征[15]。在非洲,大型食草动物的存在维持了开阔的环境,周期性火灾确保了植被再生。而在世界其他地区,维持开阔环境的主要原因是火灾。
虽然萨王纳发生火灾最初都是天然原因(如雷暴时的闪电),但后来火灾发生的模式长期受到了人类活动的影响(在非洲至少有30万年的历史),人类的演化导致火灾面积增加[12]。然而,人类活动对火灾产生实质性影响的时间相对较短,大致可以追溯到4000年前,畜牧业的发展导致火灾面积减少,因为放牧减少了地表生物燃料的数量[12]。
灰烬形式的养分、地下芽的存在、厚树皮对芽的保护以及植物有用于储存养分的大型地下器官,这些条件使植被(包括木本以及草本植物)在火灾之后能迅速恢复生长[5](图7)。
另一方面,近几十年来,人类活动极大地改变了火灾的模式,包括频率和强度。尽管区域之间存在差异,但是总体上火灾面积增加,同时每次过火区域的大小也发生了变化[12]。许多地方的政府(如巴西、博茨瓦纳、津巴布韦、南非)制定了防火和灭火方案,力图尽可能处理和扑灭所有火灾。
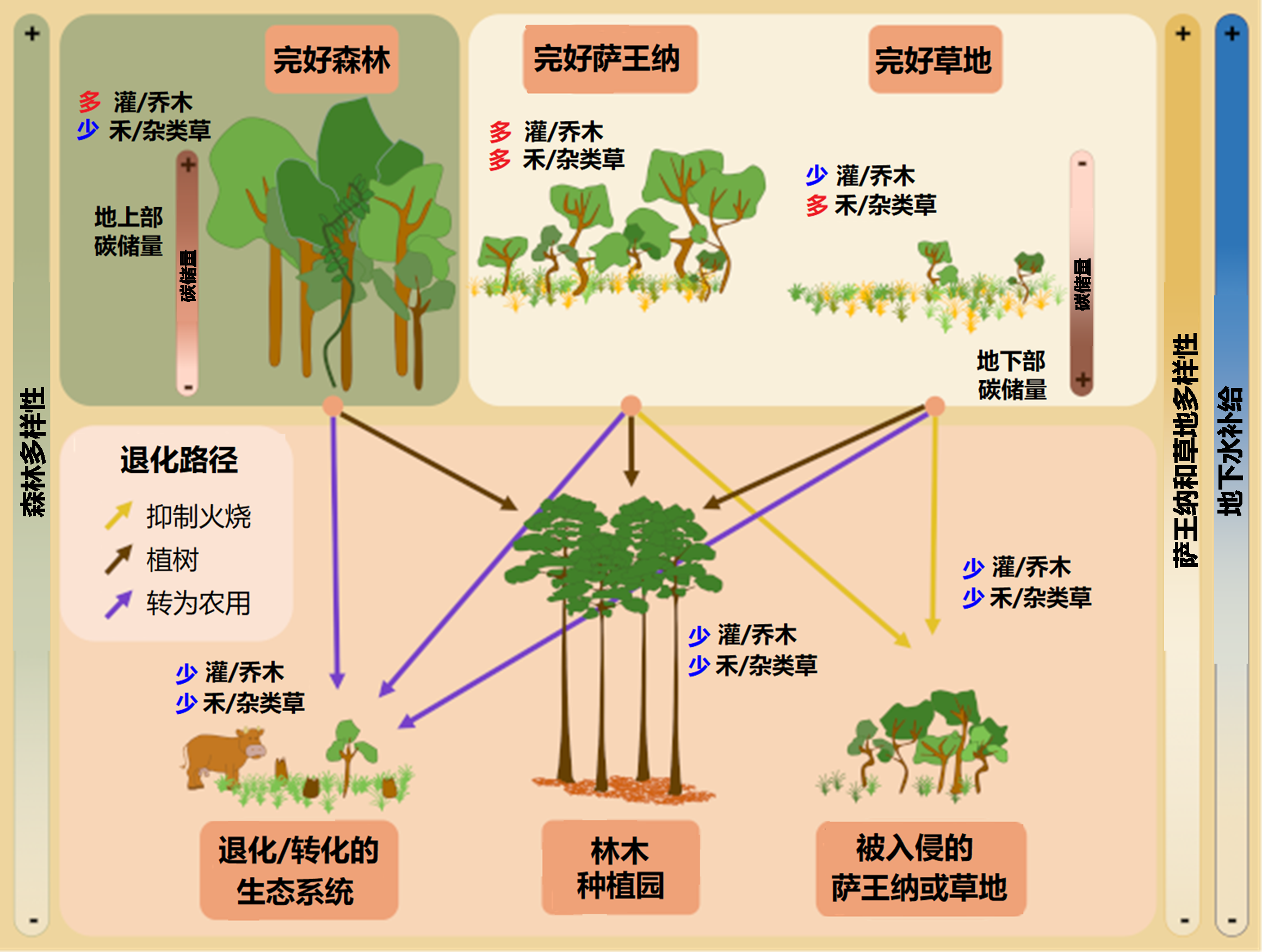
萨王纳的退化和转化可能发生得非常迅速,而且往往不可逆。退化过程多种多样,如外来物种的引入、家畜放牧、排除火灾、树木植被密集化,以及在开放环境植树造林。退化程度取决于退化的持续时间和强度。如果干扰强度大,或者发生重大环境变化,如植树造林、农田改造或者采矿等,植被和土壤的性质就会发生大幅变化,萨王纳的自然恢复能力会降低,甚至完全消失[15]。
由于以下原因,上述退化过程通常会很快超过退化阈值,使得萨王纳无法再实现自我恢复(或者需要非常长的时间):
- 成熟群落的形成依赖于许多过程的相互作用,非常缓慢;
- 恢复需要的种子传播范围往往有限。
在这些情况下,采取积极的恢复措施至关重要。
5. 恢复热带草地植物生态系统

由于萨王纳生态系统是几百万年进化和复杂生态过程相互作用的结果,因此如何恢复至今仍然是一个巨大的挑战。
第一步是要考虑火灾和大型食草动物的自然干扰,这是萨王纳形成和维持的必要条件。因此,萨王纳恢复技术要包括重新引入自然干扰,如计划烧除、放牧管理、重新引入食草动物等,此外还要清除入侵物种。
在退化严重的情况下(图8),还需要尽可能恢复地形地貌和土壤性质,然后重新引入本地物种。当然,这对于大多数缺乏生态学和生物学特征记录的草本植物来说比较困难。例如,某些物种:
- 很少产生种子甚至没有种子,或繁殖期不固定;
- 依赖火烧才能有性繁殖,或需要打破休眠状态[16]。
虽然重建草本层是恢复此类生态系统生态过程的基础,但往往难以成功,通常受到以下方面的制约:
- 入侵物种的定殖和竞争;
- 无法获得本地草本植物的种子,或者种子的繁殖效率低下[15]。
6. 植树造林
恢复萨王纳的另一个问题是它们经常被误认为是退化的森林,公众和决策者,甚至包括可能实施保护项目的政府部门,并不总能意识到萨王纳的生物多样性及其提供的(生态系统)服务的重要性。
萨王纳通常被错误地认为是退化的森林,这使人们对此类生态系统的看法复杂化,并为烧除措施提供了理由。
因为上述误解,许多所谓“恢复萨王纳”的项目都包括植树。大规模植树并不是恢复萨王纳合理技术,因为草本植物不能在荫蔽环境中生长,尤其是C4草本植物。如果出现大量树木的荫蔽,草本层的自发再生即使不是完全不可能,也会受到强烈的抑制。
在缺乏草本层生物质(食草动物的食物来源和火灾的燃料)的情况下,很难恢复接近自然状态的火灾模式、食草动物群落以及它们承担的生态过程。恢复草本层是恢复此类生态系统的首要任务。
“波恩挑战[17]”于2011年启动,目标是到2020年恢复1.5亿公顷退化土地和被砍伐的森林,到2030年恢复3.5亿公顷退化土地和被砍伐的森林。在这一背景下,出现了几项促进再造林和大规模植树的倡议,作为应对气候变化的举措,因为树木在生长过程中会吸收和储存人类活动排放的大部分二氧化碳。若干项倡议指明了可开展森林恢复的潜在区域[18],[19]。有时,植树有非常务实的目标,如防治荒漠化的有害影响和阻止沙漠推进(详见《绿色长城:绿化萨赫勒的希望?》)。在这种特殊情况下,树木提供了不可或缺的生态系统服务功能,一方面有助于防治荒漠化,另一方面当地居民高度依赖植物的多样性。虽然在恢复退化森林或实现防治荒漠化的具体目标时,这类项目的可能受到支持,但在以前的萨王纳草原以及其他热带草原地区造林导致风险必须受到谴责。
几项研究已经表明,在“开放”的生态系统中植树存在风险(即使该生态系统的树冠比较高)。萨王纳就是一个典型案例,其草本层以原始、不耐荫物种为主[1],[5]。
这里我们必须明确指出:在萨王纳这样的生态系统中植树并不是恢复,特别是外来树种大面积种植的政策方向是尤为更严重的错误[20]。同时需要谨记,大多数大规模植树项目是基于当前的气候条件而提出。然而,由于气候条变化,预计许多现在有利于树木生长的区域几年之后可能就不适合其生长,因此,在执行恢复项目时必须考虑到潜在的气候变化。
区分自然形成的萨王纳和退化的森林非常重要,天然萨王纳物种丰富,应当得到保护;而退化的森林有时被误以为是萨王纳或衍生的萨王纳,但是其中物种匮乏并且可以恢复成森林[21]。
此外,如果植树的主要理由是这是应对气候变化的最佳解决方案之一,那么同样需要重视的一点是萨王纳的大部分生物量在地下,约占总生物量的70%。因此,位于地下的碳储量绝对不可忽视,特别是因为其免受火灾影响且能保持稳定碳汇。
在这类开阔生态系统植树造林,除了导致植物和动物生物多样性丧失[22],还会造成多方面的经济损失。特别是对于特定人群,他们的放牧、狩猎等活动有赖于萨王纳,同时依赖萨王纳的供水等生态系统服务功能。除了生物多样性和动物栖息地丧失,在容易起火的生态系统中广泛植树还会增加大型林火的风险,最终导致碳储存损失(图9)。
7. 萨王纳和气候变化:相反的预测
萨王纳已经受到人为土地利用的严重影响,预计还会因气候变化和大气中二氧化碳浓度上升而发生显著变化[23]。事实上,萨王纳的分布范围取决于降水量和气候的季节性。对于未来的预测,不同的模型给出了截然不同的结果[24]。总体而言,气候变化会通过两种不同的机制减少全球萨王纳的分布范围:
- 在降水量预计会减少的地区,萨王纳面临沙漠化的风险。因为构成萨王纳的两种植被——C3树木和C4草本植物——对相同的环境因子有不同的反应[25],因此,萨王纳对干旱的响应可能不同于森林和草原[26]。此外,森林等不适应火灾的生态系统中,干旱的加剧会影响树木的生存,同时促进火灾的发生。
- 在其他降水量预测会增加的地区,乔木和灌木覆盖更加浓密(竞争干扰),危害萨王纳。由于大气中二氧化碳浓度增加,这一过程已经开始了[27]。在一些依赖畜牧业的地区,草本植物的减少可能会对当地经济造成重创。
例如,气候变化将导致塞拉多生态群落的严重同质化[28],这是生物多样性丧失和生态系统服务功能受损的主要驱动因素之一。
8. 重要信息
- 恢复退化的森林——包括重新种植树木——很重要,但是植树不应妨碍其他生态系统的保护和恢复。
- 生态恢复应恢复退化的生态系统,而不是破坏自然生态系统。
- 由于恢复萨王纳仍然很困难,因此在环境政策方面应该优先保护。
- 重视生物多样性和认识到萨王纳提供的生态系统服务功能是改善其保护、优化自然区域管理的第一步,这意味着需要运用计划烧除、允许自然火灾存在、引入本土大型食草动物等措施。
本文是勒·斯特拉迪奇和依·布韦松的“尊重”R90的修改版本,由法国生态与进化协会(SFE2)出版,并于2020年2月发布在其网站上。
参考资料及说明
封面图片:[图片来源: ©索·勒斯特拉迪奇]
[1] Veldman, J.W., Buisson, E., Durigan, G., Fernandes, G.W., Le Stradic, S., Mahy, G., Negreiros, D., Overbeck, G.E., Veldman, R.G., Zaloumis, N.P., Putz, F.E., & Bond, W.J. 2015. Toward an old-growth concept for grasslands, savannas, and woodlands. Frontiers in Ecology and the Environment 13: 154-162.
[2] Pausas, J.G., & Bond, W.J. 2020. Alternative Biome States in Terrestrial Ecosystems. Trends in Plant Science. doi: 10.1016/d.tplants.2019.11.003.
[3] Hoffmann, W.A. 1998. Fire and population dynamics of woody plants in a neotropical savanna: matrix model projections. Ecology 80: 1354-1369.
[4] Sankaran, M., Hanan, N.P., Scholes, R.J., Ratnam, J., Augustine, D.J., Cade, B.S., Gignoux, J., Higgins, S.I., Le Roux, X., Ludwig, F., Ardo, J., Banyikwa, F., Bronn, A., Bucini, G., Caylor, K.K., Coughenour, M.B., Diouf, A., Ekaya, W., Feral, C.J., February, E.C., Frost, P.G.H., Hiernaux, P., Hrabar, H., Metzger, K.L., Prins, H.H.T., Ringrose, S., Sea, W., Tews, J., Worden, J., & Zambatis, N. 2005. Determinants of woody cover in African savannas. Nature 438: 846-849.
[5] Staver, A.C., Archibald, S., & Levin, S.A. 2011. The global extent and determinants of savanna and forest as alternative biome states. Science 334: 230-232.
[6] Lehmann, C.E.R., Anderson, T.M., Sankaran, M., Higgins, S.I., Archibald, S., Hoffmann, W.A., Hanan, N.P., Williams, R.J., Fensham, R.J., Felfili, J., Hutley, L.B., Ratnam, J., San Jose, J., Montes, R., Franklin, D., Russell-Smith, J., Ryan, C.M., Durigan, G., Hiernaux, P., Haidar, R., Bowman, D.M.J.S., & Bond, W.J. 2014. Savanna vegetation-fire-climate relationships differ among continents. Science 343: 548-552.
[7] Dantas, V. de L., Hirota, M., Oliveira, R.S., & Pausas, J.G. 2016. Disturbance maintains alternative biome states (M. Rejmanek, Ed.). Ecology Letters 19: 12-19.
[8] Vasconcelos, T.N.C., Alcantara, S., Andrino, C.O., Forest, F., Reginato, M., Simon, M.F., & Pirani, J.R. 2020. Fast diversification through a mosaic of evolutionary histories characterizes the endemic flora of ancient Neotropical mountains. Proceedings of the Royal Society B: Biological Sciences 287: 20192933.
[9] Simon, M.F., Grether, R., Queiroz, L.P. De, Skema, C., Pennington, R.T., & Hughes, C.E. 2009. Recent assembly of the Cerrado, a neotropical plant diversity hotspot, by in situ evolution of adaptations to fire. Proceedings of the National Academy of Sciences of the United States of America 106: 20359-20364.
[10] Charles-Dominique, T., Beckett, H., Midgley, G.F., & Bond, W.J. 2015. Bud protection: a key trait for species sorting in a forest-savanna mosaic. New phytologist. doi: 10.1111/nph.13406; Charles-Dominique, T., Midgley, G.F., & Bond, W.J. 2017. Fire frequency filters species by bark traits in a savanna-forest mosaic (S. Scheiner, Ed.). Journal of Vegetation Science 28: 728-735.
[11] Bond, W.J., Woodward, F.I., & Midgley, G.F. 2004. The global distribution of ecosystems in a world without fire. New Phytologist 165: 525-538.
[12] Archibald, S., Lehmann, C.E.R., Gomez-Dans, J.L., & Bradstock, R.A. 2013. Defining pyromes and global syndromes of fire regimes. Proceedings of the National Academy of Sciences 110: 6442-6447.
[13] Beuchle, R., Grecchi, R.C., Shimabukuro, Y.E., Seliger, R., Eva, H.D., Sano, E., & Achard, F. 2015. Land cover changes in the Brazilian Cerrado and Caatinga biomes from 1990 to 2010 based on a systematic remote sensing sampling approach. Applied Geography 58: 116-127.
[14] Overbeck, G.E., Müller, S.C., Fidelis, A., Pfadenhauer, J. Pillar, V.D., Blanco, C.C., Boldrini, I.I., Both, R., & Forneck, E. (2007). Brazil’s neglected biome: The South Brazilian Campos. Perspectives in Plant Ecology, Evolution and Systematics. 9. 101-116. 10.1016/j.ppees.2007.07.005.
[15] Buisson, E., Le Stradic, S., Silveira, F.A.O., Durigan, G., Overbeck, G.E., Fidelis, A., Fernandes, G.W., Bond, W.J., Hermann, J., Mahy, G., Alvarado, S.T., Zaloumis, N.P., & Veldman, J.W. 2019. Resilience and restoration of tropical and subtropical grasslands, savannas, and grassy woodlands. Biological Reviews 94: 590-609.
[16] Dayrell, R.L.C., Garcia, Q.S., Negreiros, D., Baskin, C.C., Baskin, J.M., & Silveira, F.A.O. 2017. Phylogeny strongly drives seed dormancy and quality in a climatically buffered hotspot for plant endemism. Annals of Botany 119: 267-277.
[17] “波恩挑战”是一项全球倡议,其目标是在2020年恢复全球1.5亿公顷被砍伐森林和退化土地,到2030年恢复3.5亿公顷的被砍伐森林和退化土地。它由德国政府和国际自然保护联盟(IUCN)于2011年启动,并在2014年联合国气候峰会的《纽约森林宣言》中得到认可和扩展。
[18] Bastin, J.-F., Finegold, Y., Garcia, C., Mollicone, D., Rezende, M., Routh, D., Zohner, C.M., & Crowther, T.W. 2019. The global tree restoration potential. Science 365: 76-79.
[19] 森林:为人类和地球维护森林;世界资源研究所。
[20] Veldman J.W., Aleman J.C., Alvarado S.T., Anderson T.M., Archibald S. et al. 2019. Comment on “The global tree restoration potential.” Science 366, Issue 6463, eaay7976; DOI: 10.1126/science.aay7976
[21] Meli, P., Holl, K.D., Rey Benayas, J.M., Jones, H.P., Jones, P.C., Montoya, D., & Moreno Mateos, D. 2017. A global review of past land use, climate, and active vs. passive restoration effects on forest recovery (S. Joseph, Ed.). PLOS ONE 12.
[22] Abreu, R.C.R., Hoffmann, W.A., Vasconcelos, H.L., Pilon, N.A., Rossatto, D.R., & Durigan, G. 2017. The biodiversity cost of carbon sequestration in tropical savannah. Science Advances 3: e1701284.
[23] Osborne C.P., Charles-Dominique T., Stevens N., Bond W.J., Midgley G. & Lehmann C.E.R. (2018) Human impacts in African savannas are mediated by plant functional traits. New Phytologist 220:10-24.
[24] Moncrieff, G.R., Scheiter, S., Langan L., Trabucco, A. & Higgins, S.I. 2016. The future distribution of the savannah biome: model-based and biogeographic contingency. Phil. Trans. R. Soc. B37120150311
[25] 随着大气中CO2浓度的增加,C3植物的光合能力可以接近C4植物(参见光合作用的碳代谢途径)。
[26] Sankaran M. (2019) Droughts and the ecological future of tropical savanna vegetation. J. Ecol. 107:1531-1549.
[27] O’Connor, T.G., Puttick, J.R. & M Hoffman, M.T. (2014) Bush encroachment in southern Africa: changes and causes, African Journal of Range & Forage Science, 31:2, 67-88, DOI: 10.2989/10220119.2014.939996
[28] Hidasi-Neto, J., Joner, D. C., Resende, F., Monteiro, L. D., Faleiro, F. V., Loyola, R. D., & Cianciaruso, M. V. (2019). Climate change will drive mammal species loss and biotic homogenization in the Cerrado Biodiversity Hotspot. Perspectives in Ecology and Conservation. doi:10.1016/j.pecon.2019.02.001
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: LE STRADIC Soizig, BUISSON Elise (2025年1月12日), 恢复热带稀树草原和热带草地生态系统, 环境百科全书,咨询于 2025年4月8日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/restoring-savannas-and-tropical-herbaceous-ecosystems/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。