





揭示光合作用的真相

什么是地球上生命最必需的东西?当然是水;但也可能是光,提升光的光合作用对生物体是有益的。然而光合作用仅利用了到达地球的太阳能的很小一部分(在最佳条件下为5%到6%,平均不到1%)。这些光能每年可从大气 CO2 中固定 1150 至 1200 亿吨碳到生物质中(请参阅 “光合作用中碳的路径”)。在地质历史时期,这一过程使得化石燃料(煤、石油、天然气)得以生成,这些化石燃料提供了我们现代社会消耗的 80%的能源(请参阅 原油:生物起源的证据)。那么光合生物是如何收集太阳能的?它们又是如何回收太阳光所含的能量的?作为叶绿体的太阳能收集器是如何工作的?
1.自养和光合作用
“1771年8月16日,我在一个透明的封闭空间中放了一株薄荷,在密闭的空气中点燃一支蜡烛燃烧直到很快熄灭。27天后,我用凸透镜将日光束聚焦到烛芯上,再次点燃了熄灭的蜡烛,它在以前无法燃烧的空气中燃烧得很好。”这就是约瑟夫· 普里斯特利(Joseph Priestley)的实验报告(参见“光合作用的一些先驱”),该实验让他发现了氧气并揭示了绿色植物新陈代谢的基础*:即光合作用。
1.1 什么是光合作用?
[背景图片来源:Reculée des Planches, Jura, France © Pierre Thomas, Planet-Terre]
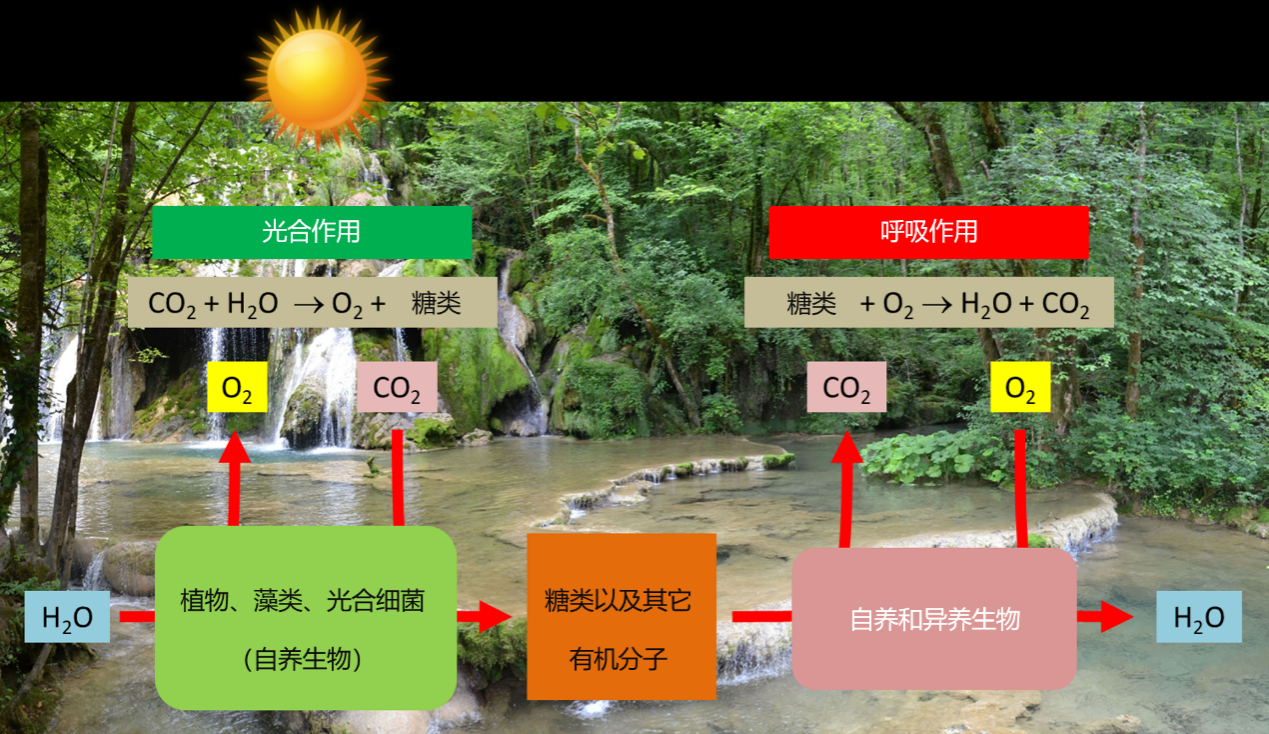
这个代谢过程为光合作用的有机体(植物、藻类、蓝藻)提供了自养的机会。由于光的作用,这些生物氧化水,释放氧气,并通过从周围环境(空气、土壤、水生环境……)中吸收矿物质合成有机物(生物质)来固定二氧化碳。以下方程式总结了该光合作用的过程(图1):
n [CO2(二氧化碳)+ H2O(水)] + 太阳能→(CH2O)n(糖)+ n O2(氧气)
蓝藻的祖先原核生物(出现在超过三十亿年前前寒武纪的原始海洋中)是最早能够进行光合作用的生物之一(参考 “生物圈,一个主要的地质参与者”)。它们利用太阳能生成氧气,使氧气在环境中缓慢积累,从而引发了一场真正的“进化革命”。原始大气中氧气的富集导致了臭氧层的形成,臭氧层可保护地球免受太阳紫外线的辐射,从而导致了气候变化和地壳成分的变化。这些变化又使细菌、动植物以新生命的形式得以在各大洲定居[1]。
1.2 叶绿体,光合作用的器官

[图片来源:雅克·乔亚德拍摄]
光合作用发生在高等植物的叶片、藻类*和单细胞微生物(藻类、蓝藻等)中。绿色植物叶子的形状通常是扁平的,它面向光线的方向,以及它薄薄的结构,使它成为太阳辐射的有效接收器(图2)。
叶细胞的细胞质中含有大量叶绿体(图3)。这些细胞器高度分化并包含所有叶绿素,专门用于完成光合作用[2]。在每克菠菜叶中,约有5亿个叶绿体。平均而言,近60%的叶片蛋白质位于叶绿体中。
- 视频“叶绿体在远端细胞中的运动”:
[图片来源:Kristian Peters — Fabelfroh / CC BY-SA 3.0]
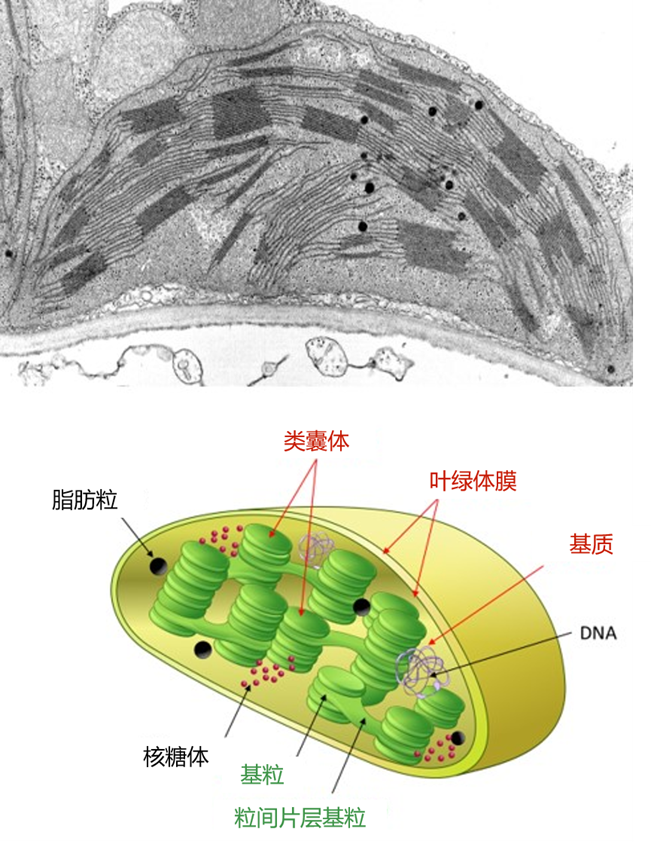
在电子显微镜下观察(图4),叶绿体类似一个卵形圆盘,长7至8微米,直径2至3微米。它包括三个部分:
- 叶绿体膜,双膜系统(由内膜和外膜组成,之间由膜间隙分隔),划分成不同的叶绿体;
- 类囊体,扁平袋状的膜网;通常堆积在基粒中,由基粒片层连接在一起。由类囊体膜包围的内部空间称为腔体。
- 叶绿体基质是受膜限制的空间,类囊体处在基质中。在电子显微镜下,它具有颗粒状外观。它包含许多酶、DNA、蛋白质合成部分(核糖体)和一些脂肪粒。
光合作用在这些不同的区室中协调发生:
- 初级反应阶段发生在类囊体内,包括光的吸收和氧的释放(参考焦点:“Z作为光合作用”);
- 然后在基质内进行二氧化碳的固定和有机碳分子的合成(参考:“光合作用的碳代谢途径”);
- 叶绿体与其细胞环境之间的分子运输涉及到叶绿体膜的作用(参考焦点:“是蔗糖还是淀粉?”)。它为生物质合成提供营养。
有关叶绿体各区室及其成分和功能的概述,可用互动方式在“SUN Chloroplast E-book”网站上查询: http: //www.markhoelzer.com/SUN-chlorophyllEbookWorking/chloroplast.html
[图片来源: 第一张图片,Eldon Newcomb拍摄,©威斯康星大学董事会;第二张图片,©叶绿体照片 [http://at-chloro.prabi.fr/at_chloro/]
2.叶片是太阳能的收集器
2.1 为什么叶子是绿色的?
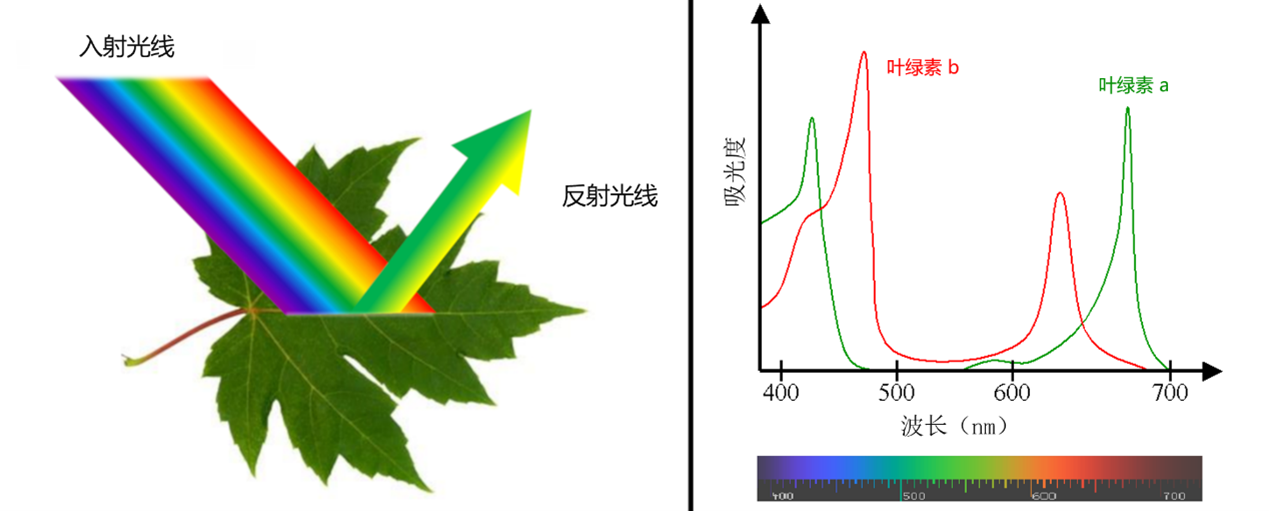
多数叶子是绿色的,少数会有白色或彩色部分。它们为什么是绿色的?这个问题的答案似乎很简单:因为它们含有叶绿素(参考: “叶片的颜色”)。
但事情要复杂一些,因为这涉及到我们的视觉和大脑协调的特殊特征。我们只看到激活我们视网膜细胞中受体的电磁光谱的波长。这些接收器对3种颜色(蓝色、绿色和红色)很敏感,并能检测环境中物体反射到眼睛里的光。它们使我们能够看到彩虹光谱中无限种深浅不一的颜色(紫色、靛蓝、蓝色、绿色、黄色、橙色、红色)[3]。但是,各种各样的色盲无法区分所有的颜色,最常见的混淆的颜色是绿色和红色。每个动物物种都有其特定的视力范围(例如蜜蜂在紫外线下才能看见),很少有动物能看到绿叶(参考: “光、视觉、生物钟”和“天空的颜色”)。
[右侧照片来源:叶绿素a/b的光谱特征,PNG: Daniele Pugliesiderivative work: M0tty / CC BY-SA 3.0].
对于光合作用,光的质量比数量更重要。例如一分子叶绿素a(最丰富的叶绿素形式)能有效地吸收紫色、红色、橙色和一些黄色的光。叶绿素b在光谱的蓝色部分更有效。另一方面,它们吸收绿光的效率非常低,所以绿光被叶片反射。这就是为什么叶子对我们来说叶片是绿色的(图5)。如果光谱所有范围地光都能被叶绿素很好地吸收,那么即使在白天植物对我们来说也会显得很黑!
2.2 叶片需要多少光?
实际上,叶片吸收的阳光不到它们接收的阳光的1%,因此,光照强度通常不是限制因素[4]。但是非常强的光会导致叶片中能量过多,从而造成光抑制:形成氧化胁迫*就能破坏光捕获结构(请参阅 “植物如何应对高原反应?”)。
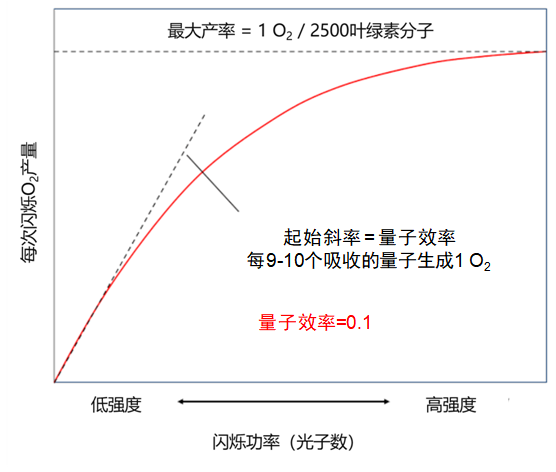
[图片来源:改编自参考文献[5]。]
光照是光合作用的驱动力。在实验中,我们观察到光合作用强度随光照强度的增加而稳定增加。但是,高强度时会发生饱和:某些反应变成了限制因素(例如CO2的浓度或温度)。这些实验观测结果揭示了两种反应的存在:一种是需要光的反应(所谓的“光”或光化学反应),另一种可能是在没有光的情况下发生的反应(不恰当地称为“暗”反应,而是生物化学反应)。
1932年,Emerson [5]和Arnold用持续数毫秒的强闪光照射小球藻(一种可进行光合作用的单细胞绿藻),并证明了闪光提供的光能可使2,500个叶绿素分子释放出一个氧气分子(图6)。大约需要吸收9到10个光子才能产生这样一个氧气分子。这对应于约0.1的量子效率,即释放的氧分子数与吸收的光子数之比。这个实验引出了光合单元的概念,稍后将以光系统的特征来说明。
2.3 所有波长的光在光合作用过程中所起的作用都一样吗?

所有波长的光对光合作用的重要性都一样吗?19世纪Engelmann [6]在一个实验中证实了光颜色的重要性。实验中用棱镜分离出不同颜色的光照射丝状藻类(螺旋类),然后用好氧细菌的相对数量作为氧气产量的指示器。细菌密度最高出现在蓝色和红色灯光照射的区域。
根据不同波长测量光合作用活性表明,光合作用在整个可见光范围内都是有效的,即使是叶绿素效率不很高的500-600 纳米范围内(见图5)。这归因于同样能够吸收光能的辅助色素。尤其是类胡萝卜素,它能吸收从紫色到红色波长范围内的所有光。因此,所有色素实际上能吸收整个波长范围内的光能,并将其释放以进行光合作用。
然而艾默生和刘易斯发现,使用单色光照射叶绿体,在波长大于680 纳米以上时显示出量子效率的急剧下降(图7)[7],而叶绿素却能够在该光谱区域继续吸收光能。这种效应被称为“红降”,表明在680纳米以上波长的光本身不能进行光合作用(这里通过氧气的释放量来测量)。另一方面,通过向暗红光中添加更短波长辐射(例如600 纳米)可以抑制这种红色下降。。这项实验表明存在两个不同的色素系统,以后将其描述为两种光系统(光系统I或PSI,以及光系统II或PSII):
- 一种是不吸收超过680 纳米的光并与氧气释放有关色素系统。
- 另一种是吸收波长超过680 纳米的光并且不释放氧气的色素系统。
这种协同效应表明,两种不同的系统在正常光照条件下协同反应,导致氧气的释放。
2.4 叶子中的色素如何组织?
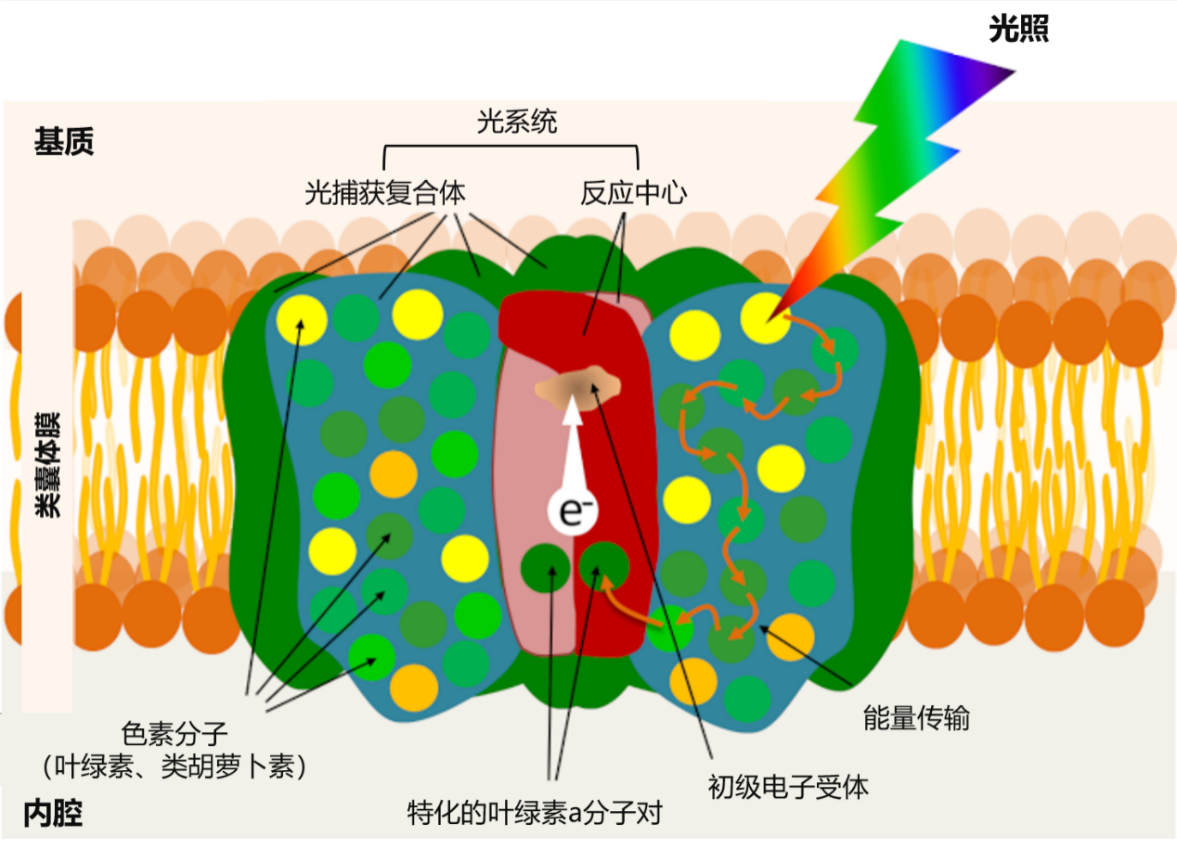
叶片的叶绿素不均匀的分布于类囊体膜内(见图5),它与光系统中的蛋白质相连,每个光系统围绕一个反应中心和一批光收集天线(图8)。这些光系统嵌入在组成类囊体膜的膜脂中。
天线结合了蛋白质和大量的感光色素:叶绿素(每个光系统中约有300个叶绿素分子),但不同的生物也会产生不同的色素,例如类胡萝卜素就存在于一些植物的光系统中。像叶绿素一样,类胡萝卜素也参与了收集天线捕捉光能的过程,然后将它们的能量转移到叶绿素。因此,光合作用首先是一种膜现象!
3.从光子到电子:光如何变成电
3.1 叶绿素和光能回收

各种天线色素(叶绿素和类胡萝卜素)主要吸收太阳辐射的可见光部分(图5),并且在捕获光子后色素分子转换为激发态(图9B)。[8] 叶绿素一旦被激发,将通过以下三个过程返回到热力学更稳定的基态(图8C):
- 通过荧光过程发射光和热(图9,C1)[9]。该机制在过量光照条件下起作用,其中过量的光能以热量的形式消散。
- 通过共振作用将能量转移到相邻分子上。色素分子之间高度接近性表明这一反应的速度极快:它在不到1皮秒的时间内发生(即不到百万分之一秒的百万分之一或10 -12秒)。这种激发能量的转移方式几乎没有能量损失。这就是天线色素(叶绿素和类胡萝卜素)如何将光带来的能量从一个分子传递到另一个分子(图9,C2),并传递到反应中心的特殊叶绿素对中。
- 通过失去电子:这就在反应中心核心内发生。
反应中心的核心是由一对特殊的叶绿素分子组成的(图7)。这种叶绿素起到能量捕获器的作用:接收由天线色素捕获的太阳光子所激发的能量。因此,天线色素所吸收的所有光能都集中在这个特殊的色素对上。
因此,被激发的“特殊”叶绿素对将电子转移到被称为初级受体的受体上(见图7),然后将其还原。这个“电荷分离”的叶绿素电子通过类囊体膜内侧运输到叶绿体基质侧。因此,反应中心可以被认为是一个分子光载体。它的正极由带正电荷的特殊氧化对形成。它的负极由带负电荷的还原分子组成:即初级受体。
还原后的一级受体很快会在一系列串联的氧化还原反应*过程中将电子传递给另一个受体,以此类推,从而使得能量从光能转化为化学能(图10)(请参考“Z作为光合作用”)。
3.2 回到最基本的问题:氧气从何而来?
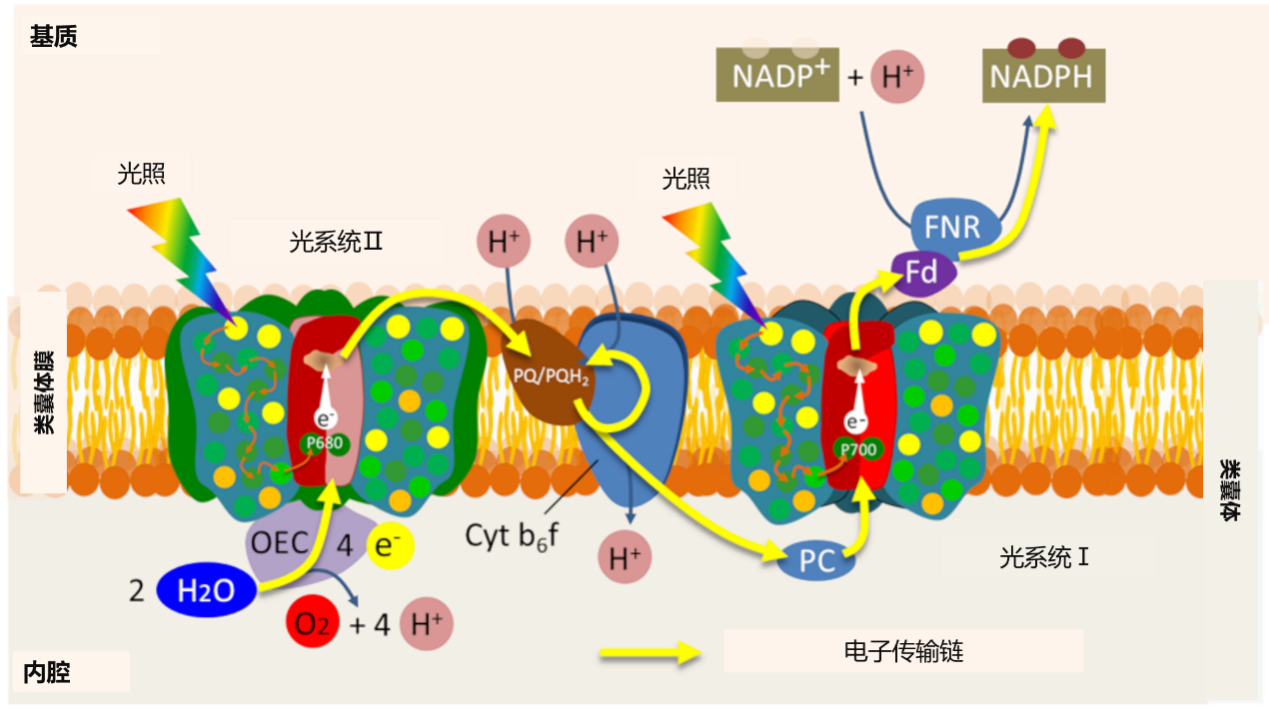
但是,为了使系统保持正常功能,反应中心中特殊的叶绿素对必须回到其基态。它们通过接受来自初级供体的电子来做到这一点。每个光系统的特征如下:
- 通过一对叶绿素吸收特定波长的光(PSI的最大值为700 纳米,PSII的最大值为680 纳米,因此其名称为P700和P680);
- 由每个光系统特定的主要供体和受体组成(图10)。

[图片来源:改编自参考文献[11]和[12]。]
在PSII中,光诱导电子离开初级电子供体,使初级电子供体P680处于氧化状态:携带一个正电荷(P680+)。为了使系统正常工作,P680+回收每个电子就至关重要。光合生物在三十亿年前进化史的早期就通过发展这一原始过程解决了这个问题:它们根据以下反应从水中获取了电子:
2 H2O(水)→O2(氧气)+4 H+(质子)+4e–(电子)(方程1)
因此,水氧化获得的电子被转移到P680+中,释放氧气,同时释放质子到类囊体腔内(图11)。这一步骤使P680+回到其中性状态(P680),并促进新的光化学循环发生[10]。
两组研究人员在1970年左右确立了这种反应机制:分别是法国的P. Joliot [11]和美国的B.Kok [12]。通过用一系列闪光照射类囊体,Joliot证明了氧气的产生具有四阶周期性,直到膜两侧差异逐渐消退(图11A)。Kok随后提供了一种解释:水的氧化循环。氧气的形成需要在PSII的供体侧连续积累四个正电荷(氧释放复合物),在该循环机制中锰扮演中心角色(图11B)。在此过程中释放的每个电子都允许一对特殊叶绿素(P680)返回其基态,并可重新获得天线色素所捕获的光子的能量。
因此我们呼吸的氧气就是该反应的副产品[13]。
3.3 电流如何流动?
在PSII的初级受体上,从P680剥离的电子通过一系列氧化还原反应流向另一个光系统(PSI),使氧化后的P700+恢复到初始状态。光系统的串联工作,将其光化学反应能量耦合到电子转移链上(参阅焦点“Z作为光合作用”)。
因此,电子转移是在光合膜内组织起来的,通过基质中的铁氧化还原酶(FNR),使得磷酸酰胺腺嘌呤二核苷酸(NADP+)化学还原。电子转移的最后阶段形成具有还原能力的NADPH。
因此,真正的电流从类囊体膜上的水(腔内)流到膜外基质一侧的NADP+(见图10)。可通过SUN Chloroplast E-book 网站http: //www.markhoelzer.com/SUN-chlorophyllEbookWorking/chloroplast.html,观看这一电子转移过程的动画形式。
3.4 质子梯度与电子传递的耦合
电子转移与跨越类囊体膜的质子梯度的建立相结合,通过以下一系列反应使得类囊体腔内相对于基质变得酸化(见图10):
- 水的氧化将质子释放到类囊体腔中(请参见方程1);
- 通过两个光系统之间的电子转移,细胞色素b6f复合物从基质中抽出质子,这些质子通过类囊体膜在管腔中积聚;
- 基质中NADPH的合成消耗质子,从而加剧了基质与内腔之间pH值的差异。
由光合膜两侧之间质子浓度的这种差异(或电化学质子梯度)产生的化学能,被膜蛋白ATP合成酶(一种真正的纳米机器)所利用,来合成三磷酸腺苷(ATP)(参阅焦点“ATP 合成”)。
总而言之,将光系统吸收的光子能量被转换为还原力(NADPH)和化学能(ATP)。
4.光化学反应与生化反应之间的耦合
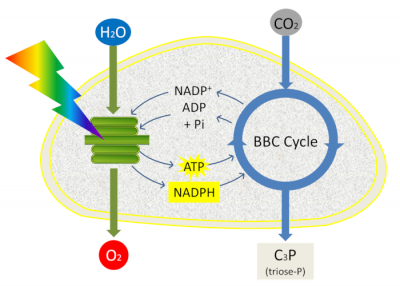
ATP和NADPH用于促进光合作用下一阶段的反应:二氧化碳的固定(参考“光合作用的碳代谢途径”)。光合作用的最后一个阶段与初级反应紧密相连(清除期),因为它使用了初级反应产生的ATP和NADPH并发生在基质中。在叶绿体基质中,通过叫做Benson-Bassham-Calvin循环中的一系列反应,将大气中CO2中的碳整合到叶绿体基质中,形成磷酸丙糖(磷酸化的3碳分子)(图12)。
然后将磷酸丙糖在叶绿体中合成淀粉、氨基酸或脂质,或从叶绿体中输出并通过细胞质酶转化为糖(蔗糖):这是所有生物体组成生物质的原料(参考焦点“是蔗糖还是淀粉?”)。
5.要记住的信息
- 自养生物在光的帮助下氧化水,释放氧气(双原子分子),并通过从周围环境中吸收矿物质合成有机物来固定二氧化碳;
- 含有叶绿体和叶绿素的植物叶片是太阳能收集器。其中叶绿素吸收蓝光和红光,但吸收绿光的效率非常低,因此绿光被叶片反射,这就是为什么叶子对我们来说是绿色的。
- 两个光系统(光系统I或PSI,光系统II或PSII)由一个集光天线和一个反应中心组成,它们通过叫做电荷分离的过程转移电子;
- 光系统通过串联将它们的光化学能量耦合到电子转移链上:因此,在光系统中接收的光子能量被转换为还原力(NADPH)和化学能(ATP)。
- ATP和NADPH用于促进光合作用下一阶段的反应:即通过发生在叶绿体基质中的Benson-Bassham-Calvin循环来固定CO2。
参考资料及说明
封面图片 [来源: © Diverticimes]
[1] Rutherford A.W. & Boussac A. (2004), Photosynthesis, a green chemistry triggered by solar energy. Photosynthesis and oxygen production. KEYS CEA 49:86-92.
[2] 1937 年,Robin Hill(罗宾·希尔,英国生物学家,1899-1991)发现叶绿体是确保光合作用的“叶绿素颗粒”。它能分离叶绿体(实际上是类囊体),并通过在人工电子受体(氧化剂)存在下悬浮发光的叶绿体来实现氧气生产。这就是“Hill反应”。
[3] 蓝光光子比红光光子具有更多能量(普朗克定律,参见“普朗克理论”)。因此,彩虹的颜色顺序对应于从红色到蓝色增加的连续能量光谱。光子的能量 (e) 由方程 e = hc/λ 给出,其中c 是光速,h是普朗克常数,λ是光的波长。爱因斯坦能量 (E) 是 E = Ne = Nhc/λ = 28,600/λ,E 单位为千卡(kcal),λ 单位为纳米 (nm; 1 纳米 = 10 -9m)。波长为 680 纳米 的红光,其爱因斯坦能量约为 42 千卡。蓝光的波长较短,因此比红光具有更多的能量。植物使用的部分太阳光谱估计平均波长为 570 纳米;因此,光合作用过程中使用的光能约为28,600/570,即 50 千卡每爱因斯坦。
[4] 即使在树冠下,光线也不是真正的限制因素。生活在那里的植物具有适应光环境的叶子结构,以平衡光捕获(光化学反应)和 CO2固定(生化反应)。植物在生长不良和扦插过程中的分布与耐荫(或不耐荫)现象有关。这种现象(称为“避光”)与光敏色素信号有关,而与叶绿素完全无关。这又是一个与光质量(即信号)有关的问题,而是与光数量(基层)关系很小。.
[5]Robert Emerson, 罗伯特艾默生 (1903-1959),美国生物学家,有许多重要成就,解释了光对光合作用的影响(量子效率、“艾默生”效应等)。他的研究首次用试验证明叶绿体中存在两个光系统。Emerson R. & Arnold W. (1932) A separation of the reactions in photosynthesis by means of intermittent light. J Gen Physiol 15:391-420.
[6] Theodor Wilhelm Engelmann ,西奥多·威廉·恩格尔曼(1843-1909),德国生理学家,在分析肌肉收缩(横纹肌)和光合作用的机制方面起到了决定性的作用。
[7] Govindjee (1963) Emerson enhancement effect and two light reactions in photosynthesis. In: Photosynthetic Mechanisms in Green Plants. Publication 1145, Published by National Academy of Sciences – National Research Council, pp. 318-334
[8] 这种激发是由于共轭键(以及离域电子)的存在:光子的到来使离域电子从基态(未激发)进入激发态。在叶绿素中,有两种激发态:高能态 (Sa) 和低能态 (Sb),具体取决于激发光子的能量(蓝色或红色)。.
[9] 植物叶绿素荧光的发射动力学是其光合性能的极好指标。因此,他的研究为准确测量干扰植物光合活性的各种胁迫的影响提供了可能。
[10] Govindjee & Coleman W. (1990) Oxygen production by plants. Special Issue For Science, January 2000
[11] Joliot, P., Barbieri, G. & Chabaud, R. (1969) A new model of photochemical centers in system-2. Photochem. Photobiol. 10, 309-329.
[12] Kok, B., Forbush, B. & McGloin, M. (1970) Cooperation of charges in photosynthetic O2 evolution-I. A linear four step mechanism. Photochem. Photobiol. 11, 457-475; Bessel Kok (1918-1979) 是荷兰裔美国生物物理学家。
[13] 大气中的氧含量在地质时期发生了巨大变化。在 25亿年之前,大气中没有氧气。从 18亿年开始,氧含量一直在 0.1% 以上。因此,在-2.5 和-1.8亿年之间氧含量有非常显著的上升,这可由当时海洋中Fe2 O3的普遍沉淀证实。目前,大气中的氧气总质量为 1,000,000 Gt,即含量为 21% (参考:“生物圈,一个主要的地质参与者”).
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: JOYARD Jacques, MOROT-GAUDRY Jean-François (2024年3月14日), 揭示光合作用的真相, 环境百科全书,咨询于 2025年4月17日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/shedding-light-on-photosynthesis/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。


