





生命对环境制约的适应

生命形式的多样性,即生物多样性,反映在表型变异(可变性状的表达)上,这些变异很大程度上是由物种内以及物种以上水平的遗传变异引起的。环境制约决定了同一物种不同种群性状的表达,最大限度地提高了个体在当地的生存和/或繁殖。随着时间的推移,人们发现了遗传变异的分子基础以及偶然性在性状进化中的重要性,从而丰富了自然选择理论。近来,诸如非遗传传递信息的跨代转移(表观遗传学)和共生(种间合作),似乎在个体和种群的适应潜力中发挥着重要作用。在达尔文去世一个半世纪之后,在人类活动引起的规模空前的第六次生物大灭绝来临之际,了解生物体如何适应快速变化环境的所有机制,显得比以往任何时候都更加必要。
1.个体差异,种群演化

这种飞蛾成天一动不动地待在白桦树树干上,捕食鸟类难以发觉(A,白体型)。19世纪英国工业化革命后,在污染地区,黑化后的黑体型(B)极为常见。
同一物种的不同个体可能遇到截然不同的环境条件。例如,生长在平原地区的植物与生长在山上的植物面临的气候制约就完全不同。同样,生活在城市、农业地区或森林地区的动物无法获得相同的资源,也不会暴露在相同的污染物中……除了这些非生物(生态系统中影响特定生物群落的物理和化学因素,与生物因素相对,构成了该生态系统生态因素的一部分。气候因素(温度、光照、空气……)、化学因素(空气气体、矿物元素……)属于非生物因素)制约因素外,每个个体还受到与其他生物体相互作用的生物(与生命有关,生态系统的生物因素是指植物群和动物群以及它们之间的关系。生命得以发展的环境。)制约因素的制约。由于环境制约是局部可变的,具有不同性状的个体将在局部被选择。适应性性状是一种形态、生理或行为特征,在特定环境中为具有该性状的个体提供生存或繁殖优势。然而,并非所有空间上的变异性状都是适应性的。
在空间变化的适应性性状中,最著名的例子是桦尺蠖(图1)。这种飞蛾整天在桦树树干上一动不动,很难被鸟类天敌发现(A,白体型)。19世纪下半叶英国工业革命期间,黑体型(图1B)成为污染地区最常见的形态,在被工业烟尘熏黑的树干上,这种黑体型比白体型更加难以发现。这种现象被称为工业黑化(一种以身体(皮肤、羽毛、毛发……)完全呈黑色为特征的动物表型。)。在这个物种中,颜色是由一个单基因决定的,该基因存在两种形式,或等位基因:白色或黑色。这是地方适应性的一个例子:在污染地区,白蛾更容易被捕食从而消失,黑蛾具有明显生存优势;在未污染地区则恰恰相反。颜色是局部选择的适应性特征;鸟类的捕食是选择因子。为了证明这一点,研究者将黑蛾和白蛾放在白色树干上,或者黑色树干上,观察两种形态在不同情况下的被捕食率。
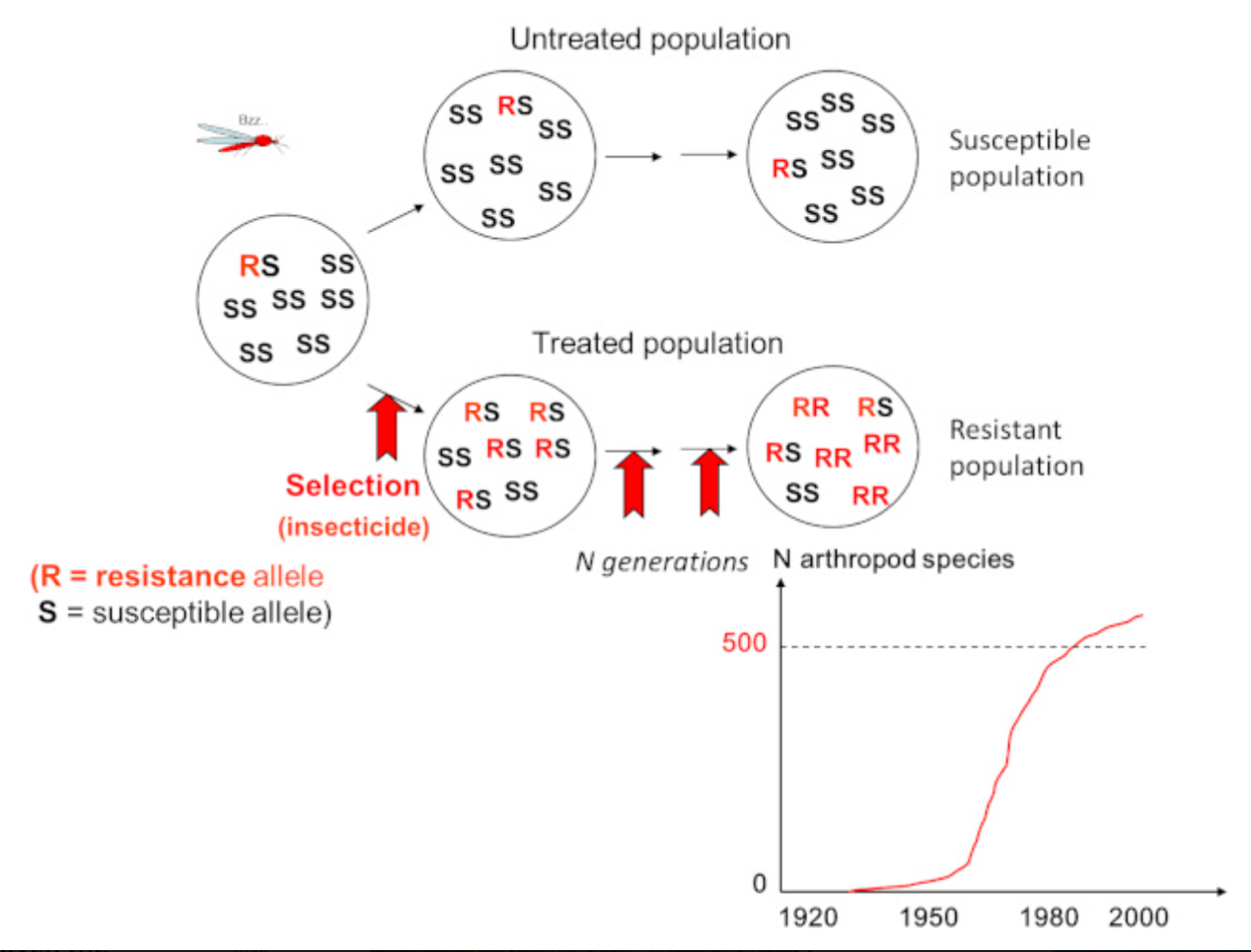
在处理过的种群中,仅少数具有抗性的昆虫存活,几代后种群就完全具有抗性。个体是选择的目标,但通过几代最具抗性个体的选择,逐渐向越来越高的抗性进化的是种群。自20世纪50年代以来,人们对病媒(蚊子)和作物害虫种群频繁使用化学杀虫剂,导致了所有处理物种中抗性种群的进化。人们不再对这些有毒分子感兴趣,因为它们已经失效了。在医院中也观察到类似的现象,抗生素的使用导致筛选出了耐药的病原菌。
当地适应的例子很多,涉及所有生物,例如:
-经杀虫剂处理的昆虫种群很快对杀虫剂产生抗药性(图2);
-医院筛选出抗生素抗药菌;
-鸟类喙的大小取决于当地可取食种子的大小[1];
-温带昆虫选择冬季滞育;
-海拔较高的植物通常比平原上生长的同种植物更小(图3);

种子分别源自海拔3000米(左侧)、1000米(右侧)处生长的母本植株。两株植物呈现完全不同的形态:左侧的植物更加紧凑,花小而少;右侧的植物则比较松散,植株较高,花更多。然而,它们栽培的条件相同(劳塔雷特高山花园,海拔2000米)。在原位(即采集种子的地方)和普通花园(进行实验的地方)分别测量这些性状,可以区分环境和基因在性状表现中所占的比例。测量原位移植个体的存活和繁殖,再与生活在初始条件下的个体相比,可以显示出这些形状与海拔高度相关的适应性。[来源:照片©皮埃尔·德·维尔梅梅鲁伊(Pierre de Villemereuil)]
在自然种群中,仅少数性状是由单个基因编码的离散性状(白/黑)(只能呈现非常明显状态的特点,例如:黑或白;有或没有智齿。)。重量、形状或大小则是种群中连续变化的性状:它们是数量性状(多基因控制);此外,这些性状的表达依赖于环境:无论携带何种基因的植物,经过充分浇水和施肥,都会比种植在干旱和贫瘠环境中的同种植物生长得大。生物体依据所处环境中的可用资源调整其性状的能力,称为表型可塑性。自然种群中的性状值(表型)部分取决于个体的基因,部分取决于个体发育的环境。
2.环境能直接影响适应性性状吗?
根据自然选择理论[1],只有性状的遗传决定部分才能遗传给后代(性状的可遗传部分)。因此,适应性性状在有利的环境中通过个体的不同生存和繁殖而逐渐增加。在当地,优势性状的固定取决于迁移个体流动的强度,它们会重新引入相反的性状特征,从而抵消当地选择的影响。然而,越来越多的最新研究表明,无论基因如何,父母所经历的环境可能会影响后代性状的表达(表观遗传学)(参见表观遗传学:基因组及其环境)。例如,研究表明,在不同氮含量的植物上饲养的毛毛虫所产生的后代,相对父母而言,在利用同类型植物方面更加高效[2]。同样,接触病原体的毛毛虫产生的后代会受到保护,使其免受病原体的侵害。卵子不仅含有双亲的基因,还含有额外的分子(成长因子、免疫因子……),这些分子可以解释这种跨代遗传。但更令人惊讶的是:当祖父经历压力(与电击、污染物相关)两代后,体外受精的小鼠仍存在对相同压力的应激反应。后代中可能存在表观遗传标记[3](由专门的酶对 DNA 或构成 DNA 的蛋白质(组蛋白)进行的生化修饰。最典型的品牌是 DNA 上的甲基(CH3),以及组蛋白的各种化学修饰(甲基化、乙酰化……)。)[3],这种标记影响应激反应相关基因的表达。这种关于当地环境特征的信息从父母直接传递给子女(后天性状的遗传[4])显然比自然选择更加有效,使个体易于适应他们可能遇到的环境条件,但这种适应现象的普遍性还远未确定,所涉及的确切机制仍有待揭示(参见 适应:应对环境挑战)。
3.个体的环境主要由其他物种构成

除了非生物环境条件(气候、污染物),每个个体都是至少一个其他物种的猎物和/或捕食者:一个有机体的环境主要是由所有其他物种组成的!与其他物种间的相互作用(见共生和寄生):根据“红皇后”(利·范·瓦伦 (Leigh Van Vale)提出的进化生物学假说,可概括如下:“一个物种必须永久进化以保持跟随与其共同进化的物种的进化的能力”。 它的名字来源于刘易斯·卡罗尔(Lewis Carroll)的书《穿越魔镜》,《爱丽丝梦游仙境》的第二部:主角和红皇后展开了一场疯狂的竞赛。 爱丽丝接着问道:“但是,红后,很奇怪,我们跑得很快,周围的风景却没有变化? ” 女王回答说:“我们跑是为了呆在同一个地方”。)假说,捕食者、寄生虫、竞争对手以及互惠者(如传粉者,图4)构成塑造种群适应性性状的主要选择因素。另一种“国王的傻瓜”(安东尼·巴诺夫斯基(Anthony Barnovsky)提出的与“红皇后”相对立的假说。该假说认为,在地质尺度上,物种的转变并不是由物种之间的竞争引起的,而是由非生物环境引起的。桦尺蠖的案例说明了“国王的傻瓜”的这一假说。)假说认为,非生物因素(陨石坠落、火山活动、气候变化)在生物多样性的演变中起重要作用。在种群和物种的进化过程中,这两种机制可能都发挥着重要作用:分支进化(一组或一群生物,其成员无论多么不同,都来自同一个共同的祖先群体:它是一个单系群体。在系统发育树中:包含祖先及其所有后代的树的分支。)(宏观进化)是渐进的(“红皇后”节奏);而有时非常迅速的物种分化(新生命物种出现的起源的进化过程,这些物种从属于原始物种的种群中个体化。)事件是与环境剧烈变化(“国王的傻瓜”)有关[5]。
4.为什么选择一些明显不利于生存的性状?

例如雄鹿的巨大鹿角,也可能与雌性的选择有关,例如孔雀鱼、孔雀或军舰鸟的艳丽色彩。
某些夸张特征的演变,如孔雀的巨大尾巴或孔雀鱼的鲜艳颜色,似乎很难解释,因为它们在生存方面似乎相当不利(更容易被捕食者发现,体型庞大……);然而,雄性动物选择这些生存代价高昂的性特征,是因为它们可以更好地接近雌性动物(通过雄性间直接竞争),或者雌性动物倾向于选择具有这些特征的雄性动物进行交配(图5)。这就是性选择[6]。
人们提出了很多个假说,来解释雌性刻意选择这种带有明显生存障碍的雄性的现象:“优质基因”假说(Good genes),认为雌性选择的性状与雄性的遗传品质有直接联系。例如,羽毛的鲜艳颜色表明雄性具有抵抗当地寄生虫的基因,这种基因将会传递给后代(汉密尔顿-祖克假说)[6]。根据“性感儿子”假说(Sexy son),选择的性状不需要与雄性的良好遗传品质联系在一起,它取悦雌性本身就是一种适应性性状,因为这种雄性的后代也会取悦雌性。根据残疾假说(Disability hypothesis),雌性依据这些特征来评估雄性的生存能力…然而这些特征却是先天降低存活率的。
5.合作与适应:全息生物论

寄主细菌的基因决定蚜虫能适应何种特定植物,而非蚜虫本身基因决定。由于细菌比蚜虫具有更高的繁殖率,这种相关伙伴的适应机制可能是生物体快速应对环境突然变化(如轮作等)的一种非常有效的方法。此外,个体的颜色是由产生绿色色素的共生细菌赋予的,通过伪装提供更好的保护来抵御捕食者(瓢虫)(见参考文献[7])。
在生命的所有尺度上,共生体无处不在(参见 共生与进化&共生和寄生)。因此,线粒体内共生(两种生物之间的互利合作,因此是一种共生形式,其中一种生物被另一种生物所包含。在细胞层面,代表了真核细胞中细胞器(线粒体和叶绿体)的形成过程。因此,线粒体是由一种细菌(可能是一种α-蛋白细菌)整合到原始真核细胞中形成的。叶绿体则是蓝藻融入真核细胞后形成的。这些转变伴随着基因从内共生体转移到宿主细胞以及新陈代谢的整合。)是真核细胞的起源:细胞代谢所必需的线粒体起源于游离细菌,可能是较大细胞的消耗物或寄生虫。共生现象存在于珊瑚礁(藻类和刺胞动物)或菌根(真菌和植物根)。我们肠道中的数十亿细菌也是如此,它们对我们的健康至关重要(参见 人类微生物群:我们健康的盟友)。在进化论中考虑共生,可以调和关于生物体适应其环境过程中先天(遗传)和后天(表观遗传,包括环境导致的可能代代相传的变化)因素所占比例的争议,并为宏观进化水平上观察到的物种形成速率变化提供新的见解。
参考文献和说明
封面照片:Grand Veymont, a group of ibexes – [Source: © Alain Herrault, www.alainherrault.com]
[1] Darwin CR (1959) On the origin of species by the means of natural selection, https://fr.wikipedia.org/wiki/De_l’origine_des_esp%C3%A8ces
[2] Cahenzli F & Erhardt A (2013) Transgenerational acclimatization in an herbivore-host plant relationship. Proc R Soc B, 280 20122856; DOI: 10.1098/rspb.2012.2856
[3] Goldberg AD, Allis CD & Bernstein E (2007) Epigenetics: a landscape takes shape. Cell 128, 635-638
[4] Sano H. (2010) Inheritance of acquired traits in plants. Plant Signaling & Behavior. 5(4):346-348
[5] Van Valen L (1977) The red queen. The American Naturalist 111(980):809-810
[6] Hamilton WD & Zuk M (1982) Heritable true fitness and bright birds: a role for parasites? Science 218, 384–387
[7] Tsuchida T, Koga R, Shibao H, Matsumoto T & Fukatsu T. (2002) Diversity and geographic distribution of secondary endosymbiotic bacteria in natural populations of the pea aphid, Acyrthosiphon pisum. Molecular Ecology 11(10):2123-2135.
[8] Arnold C. (2013) The hologenome: A new view of evolution. New Scientist 217(2899):30-34.
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: DESPRÉS Laurence (2024年3月12日), 生命对环境制约的适应, 环境百科全书,咨询于 2025年4月1日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/the-adaptation-of-life-to-environmental-constraints/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。
