



L’adaptation des organismes à leur environnement
PDF
La diversité du vivant, ou biodiversité, se traduit par des variations phénotypiques (expression de traits variables) elles mêmes causées en grande partie par des variations génétiques aussi bien à l’intérieur des espèces qu’au niveau supérieur ou supra-spécifique. Les contraintes environnementales rencontrées par les différentes populations d’une même espèce façonnent l’expression des traits maximisant la survie et/ou la reproduction des individus localement. La théorie de la sélection naturelle a été enrichie au fil du temps par la découverte des bases moléculaires de la variation génétique et de l’importance du hasard dans l’évolution des traits. Plus récemment d’autres mécanismes tels que le transfert non génétique d’information d’une génération à l’autre (épigénétique) et la symbiose (coopération interspécifique) semblent avoir un rôle important dans le potentiel adaptatif des individus et des populations. Plus d’un siècle et demi après Darwin et à l’aube d’une sixième extinction de masse d’une ampleur inédite initiée par les activités humaines, il est plus que jamais nécessaire de comprendre tous les mécanismes à l’œuvre dans l’adaptation des organismes à un environnement en pleine mutation.
- 1. Les individus diffèrent, les populations évoluent
- 2. L’environnement peut-il influencer directement les traits adaptatifs ?
- 3. L’environnement, c’est avant tout les autres !
- 4. Pourquoi certains traits apparemment défavorables à la survie sont-ils sélectionnés ?
- 5. Coopération et adaptation : la théorie de l’holobionte
1. Les individus diffèrent, les populations évoluent

Les différents individus d’une même espèce peuvent rencontrer des conditions environnementales très variables. Ainsi, une plante poussant en plaine ne subit pas les mêmes contraintes climatiques qu’une plante poussant en montagne. De même, un animal vivant en zone urbaine, agricole ou en forêt n’aura pas accès aux mêmes ressources et ne sera pas exposé aux mêmes polluants… On parle alors de contraintes abiotiquesFacteurs physico-chimiques d’un écosystème influençant sur une biocénose donnée. Opposables aux facteurs biotiques, ils constituent une partie des facteurs écologiques de cet écosystème. Les facteurs climatiques (température, lumière, air..), chimiques (gaz de l’air, éléments minéraux..) sont des facteurs abiotiques., par opposition aux contraintes biotiquesAyant un rapport avec la vie. Les facteurs biotiques d’un écosystème sont la flore et la faune ainsi que les relations qu’ils ont entre eux. Milieu dans lequel la vie peut se développer., liées aux interactions entre organismes vivants. Les contraintes environnementales étant localement variables, des individus présentant des traits différents vont être sélectionnés localement. Un trait adaptatif est une caractéristique morphologique, physiologique, ou comportementale, qui procure un avantage de survie ou de reproduction aux individus qui présentent ce caractère, dans un environnement donné. Cependant, tous les traits variables dans l’espace ne sont pas nécessairement adaptatifs.
L’exemple le plus connu d’un trait adaptatif qui varie dans l’espace est celui de la phalène du bouleau, Biston betularia (Figure 1). Ce papillon nocturne passe la journée immobile sur les troncs de bouleau, invisible aux oiseaux prédateurs (A, forme blanche typica). Lors de la révolution industrielle de la seconde moitié du 19ème siècle en Angleterre, la forme carbonaria (B) est devenue majoritaire dans les zones polluées, étant moins visible que la forme blanche typica sur les troncs noircis par les fumées industrielles. On parle alors de mélanismePhénotype animal caractérisé par la couleur entièrement noire du corps (peau, plumes, poils…). industriel (Figure 1). Chez cette espèce, la couleur est déterminée par un seul gène qui existe sous deux formes, ou allèles : blanc ou noir. Il s’agit d’un exemple d’adaptation locale : les papillons blancs sont plus consommés dans les régions polluées et disparaissent au profit des papillons noirs, et inversement en zone non polluée. La couleur est le trait adaptatif localement sélectionné, le facteur sélectif est la prédation aviaire. Pour le démontrer, on place des papillons noirs et blancs sur des troncs blancs, ou sur des troncs noirs, et on observe le taux de prédation sur les deux formes dans chacune des situations.
Les exemples d’adaptation locale sont très nombreux et concernent tous les organismes vivants, on peut citer :
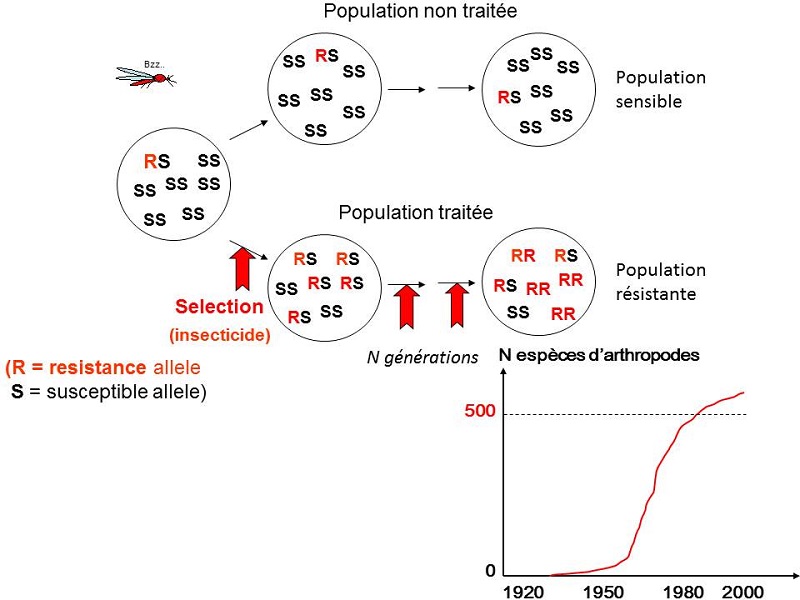
– les populations d’insectes traitées aux insecticides deviennent rapidement résistantes à ces insecticides (Figure 2) ;
– des bactéries résistantes aux antibiotiques sont sélectionnées en milieu hospitalier ;
– la taille des becs des pinsons varie selon la taille des graines disponibles localement [1] ;
– la diapause hivernale est sélectionnée chez les insectes en climat tempéré ;
– les plantes en altitude sont généralement plus petites que celles de la même espèce qui poussent en plaine (Figure 3).

2. L’environnement peut-il influencer directement les traits adaptatifs ?
D’après la théorie de la sélection naturelle [1], seule la part génétiquement déterminée d’un caractère est transmise à la descendance (part héritable du caractère). Les caractères adaptatifs augmentent donc graduellement en fréquence dans les environnements qui leur sont favorables via la survie et la reproduction différentielles des individus. Localement, la fixation d’un trait avantageux dépend de l’intensité du flux de migrants qui viennent contrecarrer l’effet de la sélection locale en ré-introduisant les traits contre-sélectionnés. Pourtant, de plus en plus d’études récentes montrent que l’environnement expérimenté par les parents pourrait avoir un effet sur les traits exprimés par les descendants, indépendamment des gènes (épigénétique) (lire L’épigénétique, le génome & son environnement). Ainsi, il a été montré que des chenilles nourries sur des plantes plus ou moins riches en azote avaient une descendance plus performante sur le type de plante utilisé par leurs parents [2]. De même, des chenilles exposées à des pathogènes produisaient des descendants protégés contre ces pathogènes. L’œuf n’est pas formé que des gènes des deux parents, mais contient des molécules supplémentaires (facteurs de croissance, facteurs immunitaires…) qui expliqueraient cette transmission entre générations. Mais il y a plus étonnant encore : deux générations après un stress (odeur associée à un choc électrique, exposition à un polluant…) subi par leur grand-père, des souris issues de fécondation in vitro sont conditionnées pour répondre à ce stress. Les marques épigénétiquesModifications biochimiques, apposées par des enzymes spécialisées sur l’ADN ou sur des protéines qui le structurent, les histones. Les marques les mieux caractérisées sont les groupements méthyle (CH3) sur l’ADN, ainsi que diverses modifications chimiques des histones (méthylation, acétylation…). modulant l’expression des gènes impliqués dans la réponse aux stress seraient présentes dans la descendance [3]. Un tel transfert d’information sur les caractéristiques environnementales locales directement de parents à enfants (héritabilité des caractères acquis [4]) est un moyen apparemment plus efficace que la sélection naturelle pour prédisposer les individus aux conditions environnementales qu’ils sont susceptibles de rencontrer, mais la généralité de ce phénomène dans l’adaptation est loin d’être établie et les mécanismes exacts demandent à être précisés (lire Adaptation : répondre aux défis de l’environnement).
3. L’environnement, c’est avant tout les autres !

Au-delà des conditions environnementales abiotiques rencontrées (climat, polluant), tout individu est la proie et/ou le prédateur d’au moins une autre espèce : l’environnement d’un organisme, c’est avant tout les autres ! Les interactions avec d’autres espèces (lire Symbiose et parasitisme) : prédateurs, parasites, compétiteurs mais aussi mutualistes (e.g. pollinisateurs, Figure 4) constitueraient l’essentiel des facteurs sélectifs façonnant les traits adaptatifs dans les populations selon l’hypothèse « de la reine rougeHypothèse de la biologie évolutive proposée par Leigh Van Valen, qui peut se résumer ainsi : « l’évolution permanente d’une espèce est nécessaire pour maintenir son aptitude suite aux évolutions des espèces avec lesquelles elle coévolue ». Elle tire son nom d’un épisode du livre de Lewis Carroll : De l’autre côté du miroir (deuxième volet d’Alice au pays des merveilles) au cours duquel le personnage principal et la Reine Rouge se lancent dans une course effrénée. Alice demande alors : « Mais, Reine Rouge, c’est étrange, nous courons vite et le paysage autour de nous ne change pas ? » Et la reine répondit : « Nous courons pour rester à la même place ». » [5]. Selon l’hypothèse alternative dite « du fou du roiHypothèse proposée par Anthony Barnovsky comme une antithèse de la « reine rouge ». Elle suggère que la transformation des espèces à l’échelle géologique est très peu induite par la compétition entre espèces mais due essentiellement au contexte abiotique. Le cas de la phalène du bouleau illustre cette hypothèse du « fou du roi ». » les facteurs abiotiques (chutes de météorites, volcanisme, changements climatiques) joueraient un rôle prépondérant dans l’évolution de la biodiversité. Les deux mécanismes sont vraisemblablement en jeu dans l’évolution des populations et des espèces, l’évolution des cladesEnsemble ou groupe d’organismes dont tous les membres, si différents soient-ils devenus, descendent d’un même groupe-ancêtre commun : c’est un groupe monophylétique. Dans un arbre phylogénétique : branche de l’arbre qui contient un ancêtre et tous ses descendants. (macroévolution) procédant par changements graduels (rythme « reine rouge ») avec parfois des évènements de spéciationProcessus évolutif à l’origine de l’apparition de nouvelles espèces vivantes qui s’individualisent à partir de populations appartenant à une espèce d’origine. très rapide en lien avec des changements brutaux de l’environnement (« fou du roi »).
4. Pourquoi certains traits apparemment défavorables à la survie sont-ils sélectionnés ?

L’évolution de certains traits extravagants comme la queue démesurée des paons ou la couleur vive des guppies peut sembler difficile à expliquer car ils semblent plutôt défavorables en terme de survie (plus visibles par les prédateurs, encombrants…) ; pourtant ces traits sexuels coûteux en terme de survie sont sélectionnés chez les mâles car ils permettent un meilleur accès au femelles, par compétition directe entre mâles, ou parce que les femelles choisissent les mâles qui présentent ces traits pour s’accoupler (Figure 5). C’est la sélection sexuelle [6].
Plusieurs hypothèses ont été avancées pour expliquer ce choix délibéré des femelles pour des mâles présentant apparemment un handicap de survie : l’hypothèse des « bons gènes » stipule qu’il y a un lien direct entre le trait choisi par les femelles et la qualité génétique du mâle. Par exemple, une couleur vive du plumage indique que le mâle possède des gènes de résistance aux parasites locaux, qui seront transmis à la descendance (hypothèse de Hamilton et Zuk). Selon l’hypothèse du « fils sexy », le trait choisi n’a pas besoin d’être associé à une bonne qualité génétique du mâle, le fait qu’il plaise aux femelles en fait un caractère adaptatif en soit, puisque les descendants de ce mâle plairont aussi aux femelles. Selon l’hypothèse du handicap, la femelle évalue la viabilité du mâle basée sur des traits qui diminuent a priori la survie…
5. Coopération et adaptation : la théorie de l’holobionte

Les symbioses sont omniprésentes dans le vivant, à toutes les échelles (lire Symbiose & évolution & Systèmes symbiotiques et parasites). Ainsi l’endosymbioseCoopération mutuellement bénéfique entre deux organismes vivants, donc une forme de symbiose, où l’un est contenu par l’autre. Au niveau cellulaire, représente les processus ayant conduit à la formation des organites (mitochondrie et chloroplaste) dans les cellules eucaryotes. Ainsi, la mitochondrie provient de l’intégration d’une bactérie, probablement une α-protéobactérie, au sein d’une cellule eucaryote primitive. Le chloroplaste a été formé par l’incorporation d’une cyanobactérie dans la cellule eucaryote. Ces transformations ont été accompagnées par les transferts de gènes des endosymbiotes dans les cellules hôtes et par une intégration des métabolismes. mitochondriale est à l’origine de la cellule eucaryote : la mitochondrie indispensable au métabolisme cellulaire était à l’origine une bactérie libre, peut-être consommée, ou parasite, de cellules plus grosses. La symbiose se retrouve au cœur des récifs coralliens (association entre une algue et un cnidaire) ou des mycorhizes (association entre un champignon et les racines d’une plante). C’est aussi le cas des milliards de bactéries que nous abritons dans nos intestins, qui se révèlent être des éléments clés de notre santé (voir Les microbiotes humains : des alliés pour notre santé). La prise en compte des symbioses dans la théorie de l’évolution pourrait réconcilier la polémique sur la part de l’inné (génétique) et de l’acquis (épigénétique, dont font partie les symbiotes-éléments de l’environnement potentiellement transmis d’une génération à l’autre) dans l’adaptation des organismes à leur environnement, et offrir un éclairage nouveau sur les changements de taux de spéciation observés à l’échelle macroévolutive.
Références et notes
Source photo de couverture : Sous le Grand Veymont, une troupe de bouquetins – Alain Herrault, www.alainherrault.com
[1] Darwin CR (1959) On the origin of species by the means of natural selection, https://fr.wikipedia.org/wiki/De_l’origine_des_esp%C3%A8ces
[2] Cahenzli F & Erhardt A (2013) Transgenerational acclimatization in an herbivore-host plant relationship. Proc R Soc B, 280
[3] Goldberg AD, Allis CD & Bernstein E (2007) Epigenetics : a landscape takes shape. Cell 128:635-638
[4] https://fr.wikipedia.org/wiki/Lamarckisme
[5] Van Valen L (1977) The red queen. The American Naturalist 111(980):809-810
[6] Hamilton WD & Zuk M (1982) Heritable true fitness and bright birds: a role for parasites? Science 218, 384–387
[7] Tsuchida T, Koga R, Shibao H, Matsumoto T & Fukatsu T. (2002) Diversity and geographic distribution of secondary endosymbiotic bacteria in natural populations of the pea aphid, Acyrthosiphon pisum. Molecular Ecology 11(10):2123-2135
[8] Arnold C. (2013) The hologenome: A new view of evolution. New Scientist 217(2899):30-34
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Pour citer cet article : DESPRÉS Laurence (26 décembre 2022), L’adaptation des organismes à leur environnement, Encyclopédie de l’Environnement. Consulté le 5 avril 2025 [en ligne ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/vivant/adaptation-organismes-environnement/.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.


