Déforestation, fragmentation et pollution des habitats, intensification de l’agriculture et de l’élevage, urbanisation, émissions massives de gaz à effet de serre dans l’atmosphère, acidification des océans, pêche industrielle intensive, transports d’espèces… Notre espèce en expansion exerce par ses multiples activités des pressions croissantes sur les écosystèmes de la planète. Comment les espèces et communautés écologiques réagissent-elles à ces pressions ? Peut-on prévoir leurs dynamiques ? Etant donné la multiplicité des espèces et de leurs interactions, il s’agira seulement ici en un premier temps d’identifier les principaux ressorts de l’adaptation des espèces et communautés aux changements de leur environnement physique et biologique. Puis de prendre en compte ces mécanismes pour comprendre et anticiper les dynamiques en cours, en cette période de changements globaux liés à l’intensification des activités humaines.
1. Les ressorts de l’adaptation des espèces au changement
On sait depuis Darwin que les espèces se transforment au fil du temps sous l’action de la sélection naturelle, qui trie les individus les plus aptes à se reproduire, ou plus exactement à transmettre leurs caractéristiques aux nouvelles générations. Ce processus d’évolution et d’adaptation permet aux espèces de répondre aux variations habituelles de leurs conditions de vie, à différentes échelles de temps.
Les recherches en écologie et évolution confirment depuis soixante ans que les espèces sont adaptées à la variabilité des conditions de vie habituellement rencontrées par les individus, d’une part, mais aussi à la fréquence relative des événements « catastrophiques » [1] rencontrés par les populations. Voyons comment.
1.1. Largeur de niche et plasticité ‘phénotypique’
Selon Hutchinson [2], toute espèce ou population peut être caractérisée par sa « niche écologique », définie comme la « place » occupée par cette espèce dans un écosystème et l’ensemble de ses exigences physiques, chimiques et biologiques à un instant donné. Produits de la sélection naturelle, ces exigences écologiques évoluent au fil des générations, avec la physiologie et le comportement des espèces. Ainsi la « largeur » de la niche écologique de chaque espèce est adaptée aux variations des conditions de vie habituellement rencontrées par les individus, dans le temps et dans l’espace (Lire Focus De la construction de niche à la destruction d’écosystèmes).
Figure 1. Peuplant estuaires, lagunes et littoraux méditerranéens, les mulets sauteurs (Liza saliens) tolèrent de larges fluctuations de salinité de l’eau. [Source : Ray eye, CC BY-SA 2.5, via Wikimedia Commons]Par exemple, les espèces de poissons et d’invertébrés pélagiques (i.e. de pleine mer), par exemple, sont souvent sténohalines parce que la salinité de l’eau au large du littoral varie très peu ; les espèces des estuaires en revanche sont nécessairement euryhalines, car adaptées à de larges fluctuations de salinité.
La sélection naturelle prépare les individus aux variations habituelles de leurs conditions de vie, durables ou récurrentes, en favorisant les modifications morphologiques, physiologiques et/ou comportementales adaptées à ces variations. Les individus font alors preuve d’une certaine adaptabilité, ou plasticité dite « phénotypique », qui peut être mise en place lors du développement de l’individu (et donc être irréversible), ou bien se poursuivre tout au long de la vie, accompagnant ou précédant les variations de l’environnement.
Parmi les adaptations phénotypiques irréversibles, citons la formation d’un casque protecteur chez les daphnies, conditionnée par la présence de poissons prédateurs dans leur habitat initial [3] ; ou le déterminisme de la caste chez les insectes sociaux (abeilles sociales, fourmis, termites, ..), dont les larves deviennent ouvriers, soldats (chez les termites) ou reproducteurs selon le régime alimentaire que leur administrent les ouvrier(e)s de la colonie.
Lorsque les variations de l’environnement sont récurrentes ou cycliques, la plasticité des individus peut être réversible. C’est ainsi que la phénologie des espèces est calée sur le rythme des variations journalières (cycle nycthéméral) et saisonnières de leurs habitats. Dans les régions tempérées, les arbres à feuilles caduques sont ainsi sensibles à la longueur des jours, ou photopériode. Chaque automne, ils perdent leurs feuilles, produisent des bourgeons et entrent en dormance pour l’hiver.
Figure 2. Les marmottes alpines (Marmota marmota) font d’amples réserves de graisses à la fin de l’été, avant de tapisser leur terrier d’herbes sèches et de s’y enfermer en famille pour y passer l’hiver. [Source : David Monniaux, CC BY-SA 3.0, via Wikimedia Commons]Ils accompagnent en cela maintes espèces animales, notamment d’insectes et de vertébrés terrestres (mammifères, reptiles et amphibiens, mais pas d’oiseaux) qui se réfugient dans un abri pour y affronter l’hiver en état de torpeur (hibernation, léthargie hivernale, diapause…). Chez les oiseaux, organismes à sang chaud incapables de basculement hypothermique, certaines espèces telles que geais, sittelles et casse-noix font des réserves de graines dès la fin de l’été, qui les nourriront l’hiver, tandis que d’autres (insectivores notamment, comme les martinets et hirondelles) font provision de graisse et migrent vers des latitudes plus clémentes, avant de revenir nicher à la belle saison.
De même, le cycle d’activité d’environ 24 heures (cycle circadien) de l’ensemble des vertébrés, rythmé par le cycle de sécrétion d’une hormone cérébrale, la mélatonine, est une adaptation à la longueur des jours – et à l’alternance jour-nuit (cycle nycthéméral) – sur notre planète.
Figure 3. Plant de tabac. Lorsque ses feuilles sont attaquées par un prédateur, à l’instar de nombreux autres végétaux, la plante de tabac se défend en synthétisant des composés phénoliques toxiques pour les herbivores. En outre, tout comme l’Aulne, l’Armoise ou l’Erable à sucre, elle émet une phéromone d’alarme (composé volatile) qui induit la même synthèse chez les plantes de la même espèce situées à proximité. [Source : Forest & Kim Starr, CC BY 3.0, via Wikimedia Commons]Cependant, les organismes ne sont pas de simples automates, programmés pour s’alimenter, se défendre et se reproduire dans un environnement aux variations prévisibles et contrôlées : la plupart sont également capables de réagir à des variations inattendues de l’environnement par un comportement adéquat, révélant une certaine plasticité comportementale.
Les plantes ne sont pas en reste. Les végétaux peuvent réagir aux variations quotidiennes, non seulement cycliques (intensité lumineuse, température…) mais aussi imprévisibles (prédation, infection, compétition…) de leur environnement local par des comportements adaptés. On peut observer par exemple l’orientation et/ou l’ouverture des fleurs en fonction de leur exposition à la lumière, la synthèse de composés toxiques anti-prédateurs, l’émission de phéromones attractives pour des espèces alliées ou de phéromones d’alarme destinées aux plantes voisines de la même espèce, en cas d’agression.
Figure 4. Les poulpes communs sont réputés pour leur propension à explorer et ‘manipuler’ de nouveaux objets, leur excellente mémoire visuelle et leurs capacités d’apprentissage. [Source : Albert Kok at Dutch Wikipedia (Original text: Albert Kok), Public domain, via Wikimedia Commons]Si elle existe chez les plantes, dénuées de véritables organes des sens, la plasticité comportementale est une propriété générale des animaux. Elle augmente avec leurs capacités d’apprentissage (capacités cognitives et pratiques), ainsi qu’avec leur socialité (complexité des groupes sociaux) et leur mobilité. Très observée chez certains invertébrés sociaux, comme l’Abeille domestique, ou non sociaux comme le Poulpe commun, elle culmine chez les mammifères et les oiseaux, dotés quant à eux de grandes capacités cognitives, de besoins énergétiques élevés et de systèmes sociaux souvent complexes [4].
La tolérance physiologique, les ajustements morphologiques ou physiologiques et la souplesse comportementale sont trois types de mécanismes permettant aux individus de réagir de manière adaptée aux variations habituelles de leur environnement. Ces mécanismes contribuent au potentiel d’adaptation des espèces aux changements de leurs habitats et conditions de vie, que ceux-ci soient causés par d’autres « acteurs »–les humains, dans le cas des changements globaux- ou par leurs propres activités (Lire Focus De la construction de niche à la destruction d’écosystèmes).
1.2. Stabilité des habitats et stratégies démogénétiques
A une échelle de temps plus large, couvrant plusieurs générations, les caractéristiques biologiques des espèces peuvent aussi évoluer en réponse à des fluctuations de l’environnement imprévisibles à l’échelle des individus.
Les mécanismes d’adaptation à long terme des espèces aux variations de leurs habitats sont non seulement démographiques mais aussi génétiques, puisqu’ils s’appuient sur la diversité et l’évolution génétique des populations. Les recherches en écologie évolutive confirment depuis les travaux pionniers de Mac Arthur et Wilson [5] dans ce domaine que les « stratégies démogénétiques » (à la fois démographiques et génétiques) des espèces sont adaptées à la fréquence relative des événements de type catastrophique, infligeant une forte mortalité aux populations.
l’une, misant sur la « quantité», de multiplication rapide des individus (dite « r », pour reproduction) et de dispersion au hasard, dans des habitats temporaires imprévisibles à l’échelle des individus ;
l’autre, misant sur la « qualité», de reproduction tardive et limitée mais compétitive (dite « K »), dans des habitats plus prévisibles et saturés.
Selon Pianka [6], les principaux traits de vie favorisés par la sélection de type « r »sont :
– la reproduction à un âge précoce, associée à une courte durée de vie et à une petitetaille,
– la fécondité élevée,
– la dispersion précoce des descendants (graines, œufs, larves…), par mobilité passive ou active.
Et les traits de vie associés à la stratégie « K » sont :
– la reproduction tardive, corrélée à une longue durée de vie et à une grande taille,
– la faible fécondité,
– les soins aux juvéniles, chez les animaux [7], [8], [9]
Par ailleurs, la diversité génétique des espèces croît avec le nombre d’individus ; elle augmente donc avec la surface des habitats favorables, avec leur connectivité, et avec le taux de croissance intrinsèque des populations.
La stratégie « r’ » en réponse aux décimations épisodiques.
Figure 6. Colonisation de nectarines –riches en nutriments- par des champignons microscopiques dits ‘moisissures’. [Source : Roger McLassus 1951 assumed (based on copyright claims)., CC BY-SA 3.0, via Wikimedia Commons]C’est la stratégie habituelle des microorganismes -tels que bactéries, protistes, champignons… et virus !- peuplant des habitats temporaires (hôtes animaux ou végétaux, habitats saisonniers, déchets organiques..), ainsi que des algues et champignons macroscopiques, aptes à coloniser rapidement tout nouvel habitat favorable.
Tirant parti de la fécondité et de la mobilité des individus, plutôt que de leurs adaptations et compétences en termes de survie locale, la stratégie « r » (de multiplication rapide et de dispersion) permet aux petites espèces prolifiques de rebondir après décimation locale. Le hasard des migrations et des mutations génétiques accélère également leur évolution et adaptation génétique face à la diversité (et à la distribution spatiale imprévisible) de leurs habitats potentiels. La « dispersion au hasard » d’un grand nombre d’individus suivant la stratégie « r » favorise la découverte et l’exploitation de nouvelles ressources (une nouvelle espèce-hôte, par exemple), et donc l’adaptation à de nouvelles conditions de vie. Ces caractéristiques augmentent la probabilité de survie à long terme des espèces (métapopulations) adaptées aux habitats instables[10].
Chez les végétaux, cette stratégie ‘r’ est celle de nombreuses plantes annuelles et graminées, qui dispersent en quantités pollens et graines sur de bonnes distances (via le vent ou le transport animal) et colonisent rapidement les habitats ouverts avant d’être broutées, piétinées, brûlées, fanées ou remplacées par d’autres espèces plus compétitives et longévives.
Cette capacité de résilience et d’évolution rapide des populations, qui augmente avec leur taux de croissance intrinsèque et leur diversité génétique, semble être un point fort des microorganismes et autres espèces prolifiques face aux actuels changements globaux. C’est ainsi qu’apparaissent épisodiquement, puis prospèrent allègrement, des souches bactériennes résistantes à certains antibiotiques, des lignées de champignons résistantes aux antifongiques, mais aussi de plantes messicoles résistantes à des herbicides, ou d’insectes « ravageurs » résistants à des insecticides (Voir le Focus Quelle stratégie démogénétique pour les abeilles ?).
La stratégie ’K’, peu adaptée au bouleversement des habitats.
Figure 8. Femelle hippopotame et son petit. Comme la plupart des grands ongulés (antilopes, mouflons, chevaux, rhinocéros, ..), les femelles hippopotames amphibies atteignent leur maturité sexuelle entre 4 et 7 ans et donnent naissance à un seul petit après une longue gestation (8 mois) – au rythme d’un petit tous les deux ou trois ans dans leur cas. [Source : Jnissa, CC BY 2.0, via Wikimedia Commons]A l’opposé de la stratégie ‘r’, la stratégie démogénétique (‘K’) est celle des mammifères longévifs et peu féconds, tels que les ongulés, les mammifères marins et les grands singes, qui élèvent au mieux un petit par an au sein de populations relativement restreintes (faible diversité génétique). Cette stratégie peut les pénaliser face aux bouleversements de leurs conditions de vie (Voir le Focus Adaptations au changement climatique sur la dynamique des mammifères). La survie des populations dépend alors de l’intensité des bouleversements locaux et régionaux, ainsi que de la plasticité physiologique et comportementale des individus.
Soulignons cependant que la fécondité des espèces sexuées varie, entre autres facteurs, avec la capacité des femelles à investir du temps et de l’énergie dans la survie et la reproduction de chacun de leurs rejetons. A défaut de soins parentaux, la sélection naturelle favorise les femelles les plus fécondes.
C’est ainsi que chez les végétaux, aux capacités parentales restreintes, la stratégie des grandes espèces longévives que sont les arbres, adaptés aux habitats stables et saturés que sont les forêts, s’accompagne d’une bonne fécondité qui augmente avec la taille et l’âge des individus. De même, peu capables de protéger leurs œufs qui se dispersent en mer, les femelles des grandes espèces de poissons pélagiques tels que thons et cabillauds investissent dans le nombre (grande fécondité) et non pas dans la survie individuelle de leurs rejetons.
D’une manière générale, la complexité des stratégies évolutives des espèces augmente avec la diversité de leurs interactions. Par exemple, malgré leur petite taille et la grande fécondité des femelles reproductrices, les espèces eusociales telles que les abeilles domestiques, les termites et les rats-taupes nus sont plutôt des stratèges ‘K’, misant sur l’élevage collectif des jeunes et la coopération intragroupe pour augmenter la prévisibilité de leur environnement et assurer la permanence de leurs colonies (Voir le Focus Adaptations à l’anthropisation des habitats).
2. Potentiel d’adaptation et changements globaux
On peut distinguer trois grands facteurs d’adaptation des espèces aux changements de leur environnement :
– leur diversité génétique, réservoir de réponses possibles face aux changements imprévisibles de l’environnement,
– leur taux de croissance « intrinsèque », qui favorise leur résilience après décimation, et
– la plasticité phénotypique (morphologique, physiologique et comportementale) des individus.
Si les espèces prolifiques et de petite taille (de type ‘r’), dans l’ensemble des groupes systématiques, réunissent souvent les deux premiers facteurs, le troisième caractérise les espèces animales dotées de grandes capacités cognitives que sont les mammifères et les oiseaux. Or ceux-ci sont pour partie des espèces ‘K’ de grande taille et peu fécondes, aux faibles effectifs, mal armées au plan « démogénétique » pour affronter des changements amples et rapides de leur environnement. Dans ces conditions, peut-on prévoir la dynamique des espèces et des communautés en réponse aux actuels changements globaux, que l’on peut caractériser par leur diversité, leur ubiquité et leur rapidité ?
2.1. Quelles principales réponses des espèces ?
Figure 9. Chevreuil à l’orée d’un bois. Glanant sa nourriture en forêt comme dans les près et champs, profitant de l’absence ou rareté des loups et autres grands prédateurs, le chevreuil est une espèce généraliste en expansion en Europe. [Source : Sylvouille at French Wikipedia, CC BY-SA 1.0, via Wikimedia Commons]Selon les caractéristiques biologiques et démographiques fondant le potentiel d’adaptation des espèces, on peut anticiper et vérifier sur le terrain :
l’expansion d’espèces généralistes (‘r’ et ‘K’, dont la nôtre !), dans l’ensemble des écosystèmes modifiés par les activités humaines – cas des sangliers, chevreuils, mésanges charbonnières, ramiers et corneilles en Europe ; cas des babouins olive en Afrique centrale… ;
la résilience d’espèces prolifiques de type ‘r’ (à grande fécondité, reproduction précoce, grande mobilité des juvéniles ou des adultes), après décimation locale ou régionale – cas des infestations épisodiques de criquets migrateurs, pyrales du maïs, campagnols terrestres et autres espèces dites « ravageurs des cultures » dans les régions de monoculture intensive ;
l’adaptation et l’évolution génétique rapide, voire la spéciation, d’espèces ‘r’ confrontées à de nouvelles conditions de vie, dans les écosystèmes modifiés – cas des nouvelles lignées bactériennes résistantes aux antibiotiques et des virus recombinants impliqués dans les épidémies de maladies infectieuses, en augmentation depuis quelques décennies [11]; cas des insectes et plantes annuelles résistant aux pesticides (Voir le Focus Adaptations à l’anthropisation des habitats) [12] ;
l’adaptation comportementale et l’évolution génétique plus lente d’espèces ‘K’ généralistes, commensales des humains, en expansion dans ces écosystèmes modifiées – ceci est attendu pour les espèces ‘K’ en expansion citées plus haut : chevreuils en Europe, babouins de savane en Afrique… ;
le déclin et la disparition de nombreuses espèces ‘K’ spécialistes, fragilisées par la fragmentation et/ou la transformation progressive de leur habitat et incapables de rebondir après décimation locale (voir le focus Les mammifères face aux changements globaux, sur la dynamique des mammifères),
la raréfaction et disparition d’une fraction d’espèces ‘r’, principalement spécialistes, confrontées à la disparition ou transformation radicale de leurs habitats et ressources (ex : invertébrés et poissons des récifs coralliens, insectes en région d’agriculture intensive) ;
2.2. Quelle dynamique des communautés et réseaux écologiques ?
Dans l’ensemble des écosystèmes modifiés par les activités humaines, l’expansion d’espèces généralistes aux dépens d’espèces spécialistes doit se traduire par une homogénéisation des faunes et flores régionales [14], [15] et un déclin du nombre total d’espèces à l’échelle mondiale (sauf diversification suffisante par spéciation dans les habitats émergents isolés, cf.[16]).
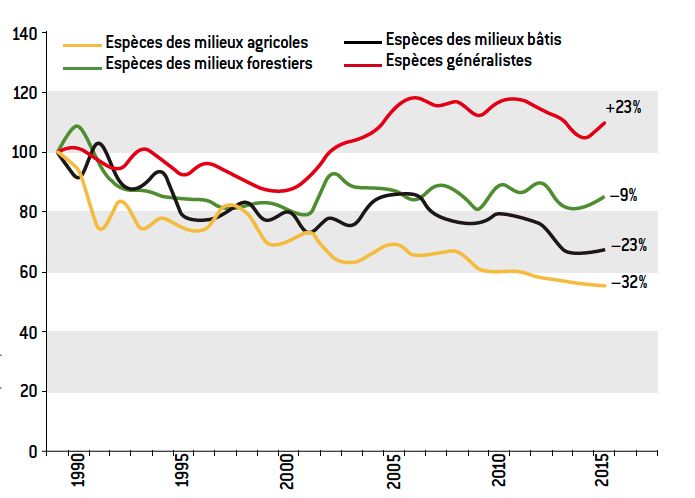
Figure 12. Suivi Temporel des Oiseaux Communs (STOC), depuis 1989 : Indicateur STOC par type d’habitat. Alors que les espèces généralistes sont en expansion en France (et ailleurs), les espèces spécialistes des habitats agricoles et autres types d’habitats sont en déclin. De ce fait, l’index de spécialisation des communautés locales d’oiseaux diminue au fil des ans, depuis plusieurs décennies. [Source : CRBPO, Vigie-Nature, MNHN]Ainsi en Europe, en réponse à l’intensification de l’agriculture, les espèces d’oiseaux généralistes (mésanges, étourneaux, corneilles noires, pigeons ramiers…), qui fréquentent également les villes, villages, parcs et jardins, composent une fraction croissante de l’avifaune des champs, tandis qu’à l’inverse les oiseaux spécialistes des milieux agricoles (linottes mélodieuses, tariers des prés, râles des genêts, huppes fasciées, vanneaux huppés…) se raréfient. Pour suivre cette dynamique de composition des communautés d’oiseaux, des chercheurs ont élaboré un index de spécialisation des communautés, qui augmente avec la perturbation des habitats [17] et confirme l’homogénéisation des peuplements d’oiseaux de France depuis 20 ans [14] et [18].
Les écosystèmes sont des systèmes organisés complexes, de résistance et résilience limitées, formés de réseaux d’espèces coadaptées, évoluant en interaction entre elles et avec leur milieu physique depuis de nombreuses générations. La modification rapide de composition des communautés, jointe aux rythmes différents d’adaptation des espèces aux changements de l’environnement, notamment du climat, désynchronisent et désorganisent ces réseaux. Ces modifications augmentent les risques de basculement des écosystèmes vers un autre état stable, défavorable à de nombreuses espèces dont souvent aux humains [19], [20] (Figure 13).
Par ses multiples activités et interactions, chaque espèce modifie son environnement et construit au fil des générations sa niche écologique.
Certaines espèces dites « ingénieures de l’environnement » ont un fort impact sur leurs habitats physiques et biologiques, et donc sur la structure et le fonctionnement des communautés locales. L’une d’elles – la nôtre! – se distingue depuis quelques siècles par la diversité et l’ampleur croissante de ses impacts sur les écosystèmes terrestres et aquatiques, actuels changements globaux.
La réponse de chaque espèce aux changements de son environnement physique et biologique dépend de facteurs externes, tels que l’amplitude et la fréquence de ces changements, et de facteurs internes, telles que sa plasticité comportementale et la fécondité des reproducteurs.
Les espèces sont adaptées à la variabilité des conditions de vie auxquelles les individus sont habituellement exposés, depuis de nombreuses générations. Leur niche écologique est donc plus ou moins large ou étroite, à l’image de ces variations : elles sont plus ou moins généralistes ou spécialistes.
En outre, elles sont adaptées à la fréquence des événements extrêmes, qui déciment les populations, par leur dynamique démographique.
On peut ainsi distinguer les espèces selon leur « stratégie démogénétique », adaptée ou non à l’ampleur et à la vitesse des changements globaux en cours.
Formés par de nombreuses espèces interagissant entre elles et avec leur habitat, les écosystèmes et réseaux écologiques sont des systèmes complexes organisés et adaptatifs, de résilience limitée.
Au delà d’un certain seuil de perturbation, les écosystèmes basculent vers un autre état, caractérisé par une autre structure (réseaux d’espèces) et un autre régime de fonctionnement.
C’est le cas actuellement pour de nombreux écosystèmes, confrontés aux changements globaux. Ces basculements en chaîne d’écosystèmes nuisent non seulement à de nombreusesespèces mais aussi aux sociétés humaines, victimes de leurs pressions croissantes sur les écosystèmes et la biosphère.
Cet article est une version revue, adaptée et actualisée de l’article « Quelles réponses des espèces et communautés écologiques aux changements globaux? » (R80a) de la même auteure, daté de juin 2018, en ligne sur la plateforme interactive de la Société Française d’Ecologie et Evolution (SFE2, https://www.sfecologie.org/regards/ ). (Regard édité par Sébastien Barot, SFE2, en février 2018).
Notes et références
Image de couverture. Pullulation de méduses communes (Aurelia aurita) sur une côte danoise [Source : Cliché Vandmaend, CC BY 3.0, via Wikimedia Commons]
[1] Evénement catastrophique, pour une population : infligeant une forte mortalité
[4] Teyssèdre A., 2006. Les clés de la Communication animale. Delachaux et Niestlé, Paris.
[5] MacArthur R.H. & E.O. Wilson, 1967. The theory of island biogeography. Princeton Univ. Press, Princeton, NJ.
[6] Pianka E.R., 1970. On r and K selection. Am. Nat. 104:592-597.
[7] Reznick D., M.J. Bryant & F. Bashey , 2002. R- and K-section revisited: The role of population regulation in life history evolution. Ecology 83(6):1509-1520.
[8] Beaumont H.J.E., Gallie J. et al., 2009. Experimental evolution of bet hedging. Nature 462:90-92.
[9] Botero C.A., F.J. Weissing, J. Wright & D.R. Rubenstein, 2015. Evolutionary tipping points in the capacity to adapt to environmental change. Proc. Natl. Acad. Sci. USA 112:184-189.
[10] Starrfelt et Kokko 2012. Bet hedging : a triple trade-off between means, variances and correlations. Biol. Rev. 87, pp. 742–755.
[13] Teyssèdre A., 2018. Les mammifères face aux changements globaux. Regards et débats sur la biodiversité, SFE, regard R80b, juin 2018.
[14] McKinney M.L. & J.L. Lockwood, 1999. Biotic homogenization: a few winners replacing many losers in the next mass extinction. T.R.E.E. 14:450-453
[15] Clavel J., Julliard R. and V. Devictor, 2010. Worldwide decline of specialist species: toward a global functional homogenization? Front. Ecol. Environ. 2:222-228.
[16] Thomas C.D., 2015. Rapid acceleration of plant speciation during the Anthropocene. Trends Ecol. Evol. 30:448-455.
[17] Julliard R., J. Clavel et al., 2006. Spatial segregation of bird specialists and generalists in bird communities. Ecol. Letters 9:1237–1244.
[18] Clavel J., 2011. L’Homogénéisation biotique. Regards et débats sur la biodiversité, SFE, regard n°16, avril 2011.
[19] Folke C., S. Carpenter et al., 2004. Regime shifts, resilience and biodiversity in ecosystem management. Ann. Rev. Ecol. Syst. 35:557-581.
[20] Cardinale B. et al., 2012. Biodiversity loss and its impact on humanity. Nature 486:59-67.
[21] Haberl H., 2007. Quantifying and mapping the human appropriationof net primary production in earth’s terrestrial ecosystems. Proc. Natl. Acad. Sci. USA 104:12944-12947.
[22] Gaston K.J., T.M. Blackburn, K. Klein Goldewijk, 2003. Habitat conversion and global avian biodiversity loss, Proc. R. Soc. Lond. B 270:1293–1300.
[23] Smil V., 2011. Harvesting the Biosphere: the Human Impact. Pop. Dev. Rev. 37(4):613-636.
[24] Teyssèdre A. & D. Couvet, 2007. Expected impact of agriculture expansion on the world avifauna. C. R. Acad. Sci. Biol. 330:247-254.
[25] Pauly D, V. Christensen V, J. Dalsgaard J, R. Froese & F.S.B. Torres, 1998. Fishing down marine food webs. Science 279:860-863.
[26] Watson R. & D. Pauly, 2001. Systematic distortions in world fisheries catch trends. Nature 414(6863):534-536.
[27] Folke C., S. Carpenter et al., 2004. Regime shifts, resilience and biodiversity in ecosystem management. Ann. Rev. Ecol. Syst. 35:557-581.
[28] Cardinale B. et al., 2012. Biodiversity loss and its impact on humanity. Nature 486:59-67.
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.
Depuis la grippe espagnole de 1918, le terme pandémie est devenu courant. Les dernières pandémies…
Emmanuel DROUET, Docteur en Pharmacie, Professeur à l’Université Grenoble-Alpes, Enseignant-Chercheur à l’Institut de Biologie Structurale UMR 5075 CNRS UGA CEA, Grenoble.