Les plantes terrestres sont régulièrement soumises à de fortes variations de température. Ces variations peuvent atteindre une amplitude de 40°C, voire plus, aussi bien dans les régions polaires que dans les zones désertiques chaudes. Enracinées, elles ont une mobilité réduite et doivent faire face aux changements qui se produisent dans leur milieu. L’assimilation du CO2 par les plantes via la photosynthèse est la porte d’entrée du carbone dans la biosphère. Quelle amplitude thermique permet son fonctionnement ? Comment réagit-t-elle aux variations rapides et lentes de la température ? Quelle est la diversité des réponses ? Quels sont les processus physiologiques qui la limitent ? Questions cruciales à replacer dans le contexte du réchauffement global.
L’accroissement actuel des émissions de gaz à effet serre va causer une augmentation de la température atmosphérique de 2 à 3 °C dans les 50 prochaines années (Lire Un cycle du carbone perturbé par les activités humaines). En même temps les vagues de chaleur et les périodes de chaleurs extrêmes vont être plus fréquentes et de plus longue durée [1]. La production agricole et le fonctionnement des forêts vont donc être très affectés. Des modèles appuyés sur des observations à grandes échelles indiquent que la baisse de rendement de nos cultures peut atteindre 17% pour chaque augmentation de 1°C de la température de la saison de croissance [2].
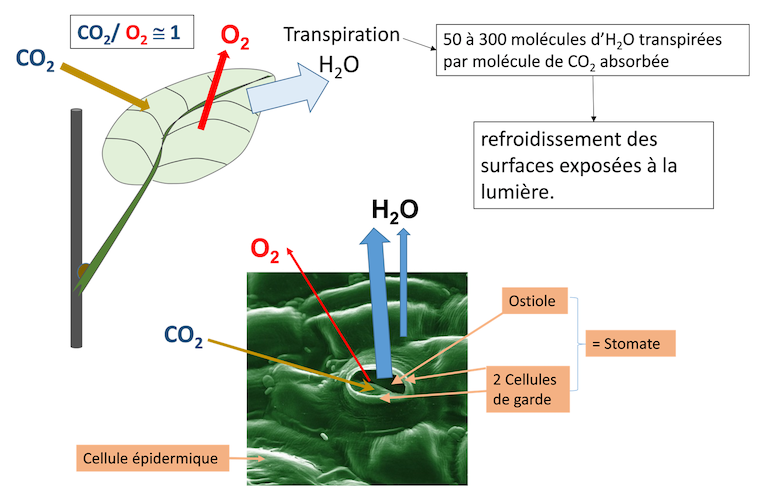
La production des plantes supérieures dépend notamment (mais pas uniquement [3]) de la photosynthèse foliaire (Lire Lumière sur la photosynthèse & Le chemin du carbone dans la photosynthèse). Le CO2 entre dans la feuille où sa réduction dans les chloroplastes est accompagnée d’une production d’O2. Son entrée se fait quasiment exclusivement par les stomates (Figure 1). Pour chaque molécule de CO2 absorbée, de 50 à 300 molécules d’eau sont transpirées au niveau des feuilles selon les plantes considérées. Cette eau permet, entre autres, le refroidissement de la feuille (voir Focus Transpiration de la feuille et protection contre la chaleur).
Figure 1. Durant la photosynthèse le CO2 absorbé et l’O2 dégagé passent principalement par l’ouverture stomatique (ostiole). La vapeur d’eau (transpiration de la feuille) passe surtout par l’ostiole mais aussi à travers l’épiderme. La transpiration permet le refroidissement de la feuille à la lumière. [Source : Schéma de l’auteur]La feuille est un convertisseur de l’énergie solaire en énergie chimique et, comme tout convertisseur d’énergie, nécessite en permanence un système de refroidissement.
Les changements climatiques qui se produisent actuellement rendent nécessaire la compréhension des effets de la température sur la photosynthèse.
2. L’optimum thermique de la photosynthèse
2.1. Schéma de la réponse thermique
L’assimilation photosynthétique de CO2 varie avec la température. Dans la majorité des cas sa réponse à la température est rapidement réversible entre 10 et 34°C environ. Dans cette gamme de températures elle présente une valeur maximum : un optimum thermique.
Figure 2. Schéma de la variation de l’assimilation de CO2 par une feuille intacte. Elle met en évidence la gamme de température dans laquelle ses variations sont généralement rapidement réversibles. [Source : Schéma de l’auteur]En dessous de 10°C et au-dessus de 34°C les plantes commencent à mettre en place des mécanismes de protection. Pour ces valeurs extrêmes, l’assimilation de CO2 est souvent instable et peut s’annuler plus ou moins rapidement : la feuille est alors soumise à contrainte (Figure 2).
2.2. Un optimum thermique calé sur la température moyenne du milieu
Les plantes de milieux froids ou dont la saison de croissance est froide ont une photosynthèse plus élevée à basses températures. Celles de milieux chauds, ou se développant durant la saison chaude, ont une photosynthèse plus élevée à hautes températures.
Figure 3. Deschampsia antarctica est l’une des deux plantes à fleurs que l’on trouve en Antarctique. Elle est souvent soumise à des températures négatives. La neige qui la recouvre fréquemment la protège contre les températures extrêmes. [Source : Lomvi2, CC BY-SA 3.0, via Wikimedia Commons]Ainsi, l’optimum thermique de l’assimilation de CO2 [4] chez Deschampsia antarctica (Figure 3) et Colobanthus quitensis, les deux seules plantes antarctiques à fleurs, se trouve entre 8 et 15°C tandis qu’il est situé vers 45°C chez Tridestomia oblongifolia, plante de désert chaud d’Amérique centrale. Cette dernière espèce détient probablement à cet égard le record du monde chez les plantes à fleurs.
2.3. L’acclimatation aux conditions thermiques du milieu
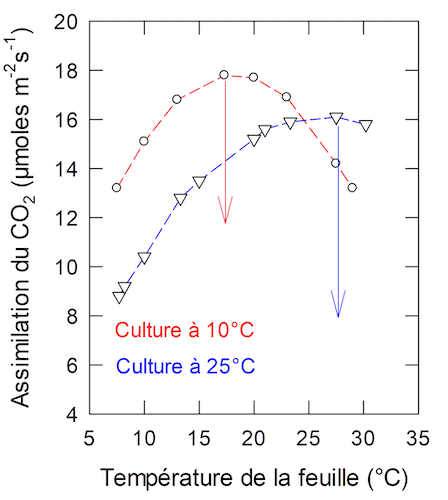
Figure 4. Variations de l’assimilation du CO2 en fonction de la température foliaire, chez une plante cultivée à 10°C (rouge) ou à 25°C. Mesures faites sur le Pois, sous une lumière proche de la saturation. Teneur en CO2 dans l’air ambiant : 390 ppm. [Source : Source : G. Cornic, résultat non publié]Des différences de la réponse thermique de la photosynthèse se trouvent aussi chez des individus d’une même espèce dont la croissance se fait à des températures contrastées. La Figure 4 montre l’assimilation du CO2 chez du Pois cultivé à 10 ou à 25°C.
Dans le premier cas (culture à 10°C) l’optimum thermique est d’environ 16°C, tandis qu’il est supérieur à 25°C dans le second (culture à 25°C). Aux basses températures l’assimilation du CO2 est plus élevée chez les plantes cultivées à 10°C.
Dans ce cas l’ajustement aux nouvelles conditions est un gain pour la plante.
De façon générale, ces changements peuvent se mesurer chez les feuilles en croissance et chez les feuilles adultes, la réponse étant de plus grande amplitude chez les feuilles en croissance.
2.5. Espèces sensibles au chaud versus espèces sensibles au froid
L’acclimatation au chaud d’espèces (ou d’écotypes [6]) adaptées aux températures fraiches se fait avec une augmentation de l’optimum thermique mais une baisse générale de la photosynthèse.
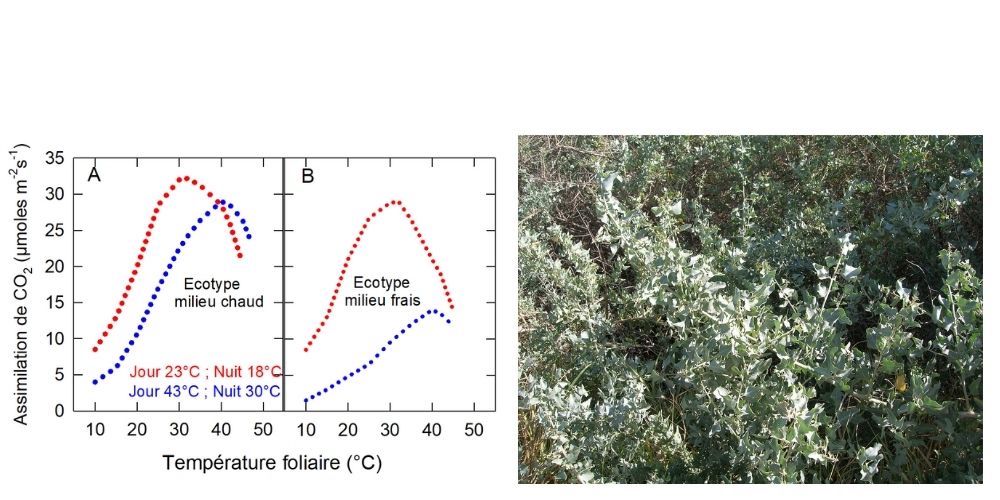
C’est, par exemple, le cas d’Atriplex sabulosa. On peut alors s’interroger sur l’intérêt de ce changement. L’inverse peut être vrai pour les plantes strictement adaptées au chaud comme Tridestomia oblongifolia. La Figure 7 illustre le cas d’Atripex lentiformis, [7] plante à feuilles pérennes, qui se trouve en Californie aussi bien dans la Vallée de la Mort que dans des habitats côtiers frais et humides :
L’assimilation de l’écotype du désert (Figure 7A) et de l’écotype côtier (Figure 7B) présentent quasiment la même réponse à la température lorsqu’ils sont cultivés sous 23°C le jour et 18°C la nuit (en rouge sur la Figure 7).
Figure 7. Variations de l’assimilation de CO2 d’écotypes d’Atriplex lentiformis venant d’un milieu chaud (A) et d’un milieu frais et humide (B). [Source : Schéma de l’auteur, d’après Pearcy (1977)] A droite, pied d’Atripex lentiformis (salt bush) [Source : Forest & Kim Starr, CC BY 3.0, via Wikimedia Commons].Sous l’alternance 43°C le jour et 30°C la nuit (en bleu sur la Figure 7), seul l’écotype du désert fait preuve de sa plasticité, conservant une assimilation de CO2 élevée dans ces nouvelles conditions. L’activité de l’écotype de la région côtière est faible à toutes les températures. Seul subsiste de ses capacités d’acclimatation le déplacement de l’optimum thermique [8].
Les plantes en C4, dont on a des traces seulement à partir de la fin de l’ère Tertiaire, ne constituent que 5% des espèces. Elles colonisent plutôt les milieux (ou les saisons) chauds et secs (Lire Restaurer les savanes et les écosystèmes herbacés tropicaux). Le Maïs et la Canne à sucre en sont des exemples.
En moyenne l’optimum thermique des plantes en C4 est situé à des températures plus élevées que celui des plantes en C3.
Cependant les plantes en C3 sont les plus plastiques. En effet leur optimum thermique varie de 7 à 35°C environ, tandis que celui des plantes en C4 oscille, à quelques exceptions près, entre 30 et 40°C. De plus, lorsque la température est inférieure à 20°C la photosynthèse des plantes en C4 est en moyenneplus faible que celle des plantes en C3.
3. L’assimilation de CO2 résulte de l’interaction de processus dont la réponse à la température est différente
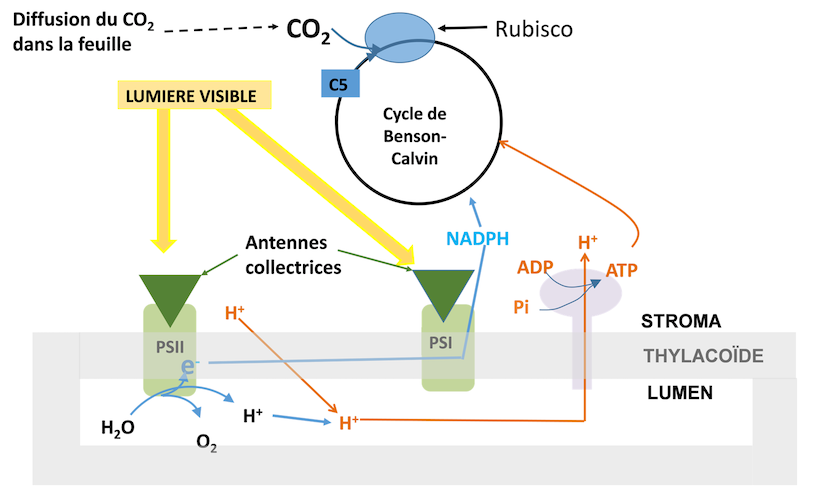
L’absorption de la lumière au niveau des antennes collectrices (Figure 9) et le transfert de son énergie vers les centres réactionnels PSII ne sont pas sensibles à la température.
Sont sensibles à la température :
La diffusion du CO2 de l’air ambiant aux chloroplastes : sa vitesse augmente avec la température.
La fixation du CO2 sur le Ribulose 1,5-bisphosphate(RuBP), un sucre dont le squelette est formé par 5 atomes de carbone. (Lire Focus Décryptage du Cycle de Benson-Bassham-Calvin)
Le transfert des électrons du PSII vers le PSI.
Figure 9. Schéma des processus en interaction lors de la fixation photosynthétique du CO2 (cas d’une plante en C3). PSI et PSII : respectivement photosystème I et II. Ils sont inclus dans la membrane du thylacoïde constituée de 2 couches lipidiques formant des « sacs » dans le chloroplaste. L’intérieur du thylacoïde est le lumen. RubisCO : enzyme qui catalyse la fixation du CO2 sur un sucre pourvu de 5 atomes de carbone (le Ribulose 1,5-bisphosphate : C5). Cycle de Benson-Calvin : permet la régénération de C5, et donne en même temps le carbone nécessaire à la plante. L’ATP est synthétisé lorsque des protons du lumen retournent au stroma à travers une ATPase en utilisant du phosphate inorganique, Pi. Les protons du lumen ont deux origines : (1) l’oxydation de l’eau dans le lumen par le PSII qui fournit aussi des électrons, e– et (2) le fonctionnement d’une pompe à protons dans le thylacoïde qui fait passer des protons du stroma dans le lumen. [Source : Schéma de l’auteur]La régénération du RuBP se fait par l’intermédiaire du fonctionnement du cycle de Benson-Calvin (C’est la « biochimie » du processus) qui utilise pour fonctionner du pouvoir réducteur (sous forme de NADPH) fournit par le transfert d’électrons. L’ATP nécessaire est synthétisé lorsque les protons accumulés dans le lumen passent dans le stroma à travers une ATPase (Figure 9).
Formation de pouvoir réducteur et synthèse d’ATP ont une sensibilité thermique voisine de celle du transfert d’électrons.
4. Quels sont les processus à l’œuvre pour fixer l’optimum thermique de l’assimilation du CO2 des plantes en C3 et en C4 ?
4.1. L’activité des Photosystèmes et du transfert d’électrons qui en résulte n’est pas en cause
Mesuré in vitro sur des thylacoïdes (voir légende Figure 9) isolés, en présence d’accepteurs artificiels, le transfert d’électrons augmente avec la température et présente un optimum thermique bien marqué. Il est situé vers 30°C et correspond à celui de l’assimilation de CO2 lorsque celui-ci est saturant[9]. L’activité du PSII a un optimum thermique identique à celle de l’activité de la chaîne de transfert d’électrons.
L’activité PSI n’est pas inhibée aux températures élevées (supérieures à 30°C, jusqu’à 45°C) où elle reste stable, voire augmente : c’est l’activité du PSII qui limite l’activité de la chaine d’électrons.
De plus le PSII est très sensible aux fortes températures qui endommagent surtout le complexe protéique qui permet l’oxydation de l’eau (voir Figure 9).
La réponse thermique du transfert d’électrons est similaire chez les plantes en C3 et en C4. Cependant il existe à cet égard, des différences d’organisation entre ces deux types de plantes (Lire Le chemin du carbone dans la photosynthèse).
La fourniture d’énergie ne peut donc expliquer les différences d’optimum thermique. C’est la façon dont l’énergie produite est utilisée qui fait la différence.
4.2. Une réponse ? Comparaison de l’effet de l’O2 atmosphérique sur l’assimilation du CO2 des plantes en C3 et en C4
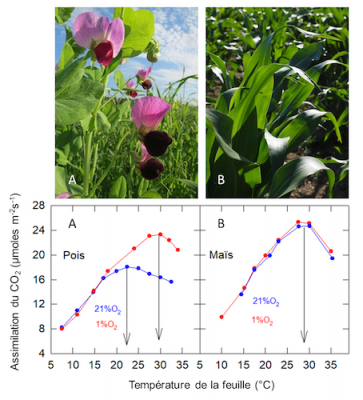
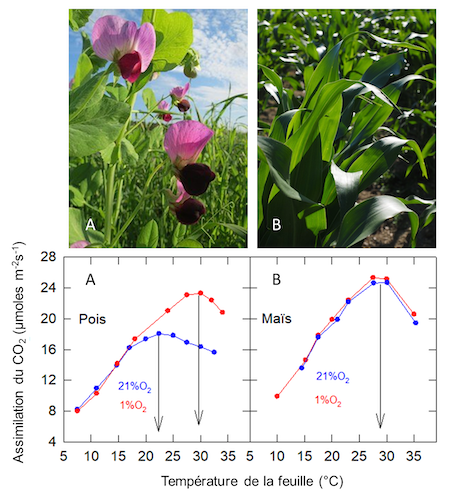
Dans un air normal[10], 21% d’O2 (+ N2) + 360 ppm de CO2: l’optimum thermique est de 27°C chez le Maïs (plante en C4), tandis qu’il n’est que de 22°C chez le Pois (plante en C3) (Figure 10) : l’optimum thermique de la plante en C4 est plus élevé que celui de la plante en C3 (voir aussi section 2.6).
Dans une atmosphère pauvre en oxygène, 1% d’O2 (+ N2) + 360 ppm CO2: l’assimilation de CO2 du Maïs n’est pas affectée, tandis que celle du Pois est stimulée au-dessus de 17°C environ, avec un déplacement de son optimum thermique qui devient voisin de celui du Maïs.
Chez les plantes en C3, l’oxygène atmosphérique inhibe l’assimilation du CO2 lorsque la température de la feuille est suffisamment élevée, alors qu’il est sans effet (ou que ces effets sont négligeables) chez les plantes en C4.
Figure 10. Variation de l’assimilation du CO2 mesurée sur des feuilles de Pois (A ; Pisum sativum) et de Maïs (B ; Zea mays) en fonction de la température foliaire. Les plantes sont cultivées en lumière naturelle à la température de 20 ± 2°C. [Source G. Cornic, résultat non publié – images de plantes libre de droits / Pixabay]Noter que la variation du transfert d’électrons estimé in vivo, en mesurant l’émission de la fluorescence chlorophyllienne en fonction de la température, est très voisine dans 1% et 21% d’O2 chez le Pois : la variation de l’optimum thermique n’est donc pas due à une modification de la photochimie.
4.3. Les propriétés de la Rubisco expliquent la différence de réponse
Cas des plantes en C3
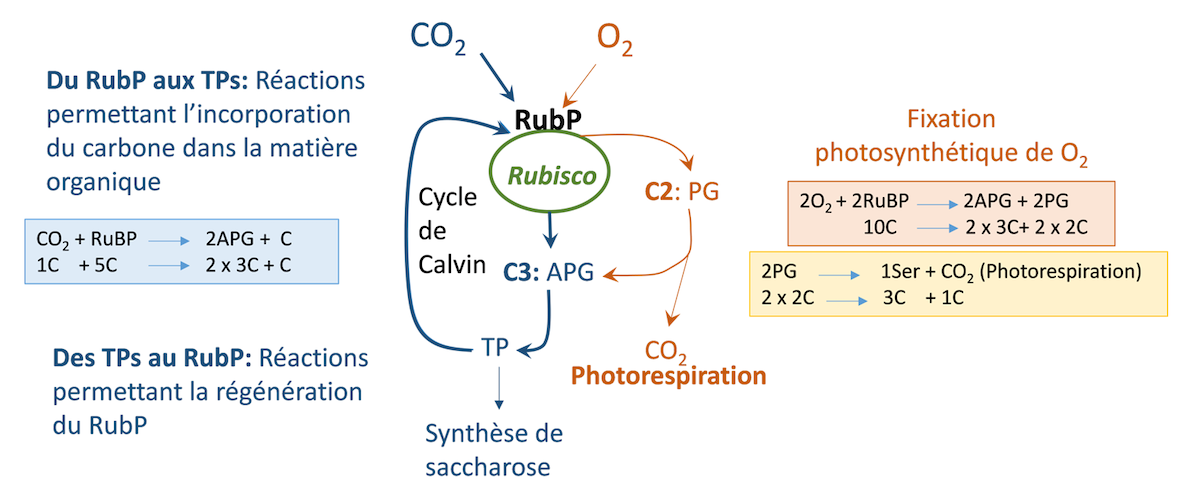
Le CO2 et l’O2 sont en compétition pour occuper les sites actifs de la Rubisco : Cette enzyme a une fonction carboxylase et une fonction oxygénase. Le CO2 entre dans le cycle de Benson-Calvin et la fixation photosynthétique de l’O2est à l’origine d’une voie métabolique responsable de la photorespiration (Figure 11 ; Lire aussi Le chemin du carbone dans la photosynthèse).
Le CO2 occupe un nombre élevé de sites actifs sur la Rubisco lorsque la teneur en O2 de l’air ambiant est faible (1% par exemple) ou celle du CO2 élevée.
L’O2 est principalement fixé si sa teneur augmenteou si celle du CO2 diminue (ce dernier libère alors des sites actifs qui sont alors occupés par l’O2).
Dans un air normal, deux raisons concourent à augmenter la fixation d’O2(et par conséquent diminuer celle du CO2) lorsque la température augmente[11].
L’affinité de la Rubisco pour le CO2 décroit plus que celle pour l’O2; facteur privilégiant l’assimilation de L’O2.
Le coefficient de solubilité dans l’eau du CO2 diminue plus que celui de l’O2 entrainant, dans le chloroplaste, une décroissance plus rapide de la quantité de CO2 que celle de l’O2 ; facteur privilégiant encore la fixation d’O2.
Dans une atmosphère pauvre en O2 (Figure 10) la compétition entre l’O2 et le CO2 est très réduite. L’énergie est utilisée alors principalement pour l’assimilation du CO2 dont la valeur augmente jusqu’aux alentours de 30°C puis décroit comme la fourniture d’énergie (voir section 4.1).
Dans un air normal, l’effet del’O2 sur la fixation photosynthétique du CO2 (Figure 11) est très faible (voir nul) lorsque la température est faible : la compétition sur les sites de carboxylation est en faveur du CO2.
Figure 11. Schéma de la fixation du CO2 et de l’O2 sur le RuBP (Ribulose 1,5-bisphosphate) chez une plante en C3. APG : Acide 3-phosphoglycérique, composé à 3 C ; TP : Trioses phosphate. Le carbone sort du cycle de Calvin pour alimenter la synthèse de saccharose. PG : phosphoglycolate, composé à 2C. Deux PG donnent une serine (Ser) contenant 3C avec la production du CO2 de la photorespiration. C = Atome de Carbone. [Source : Schéma de l’auteur]En revanche, lorsque la température augmente, la compétition sur ces sites favorise la fixation d’O2 qui consomme alors une partie croissante de l’énergie produite par l’activité des photosystèmes. Cette énergie n’est donc plus disponible pour la fixation du CO2 qui atteint sa valeur maximum vers 22°C.
Cas des plantes en C4.
Le CO2 est concentré au niveau de la Rubisco par un mécanisme insensible à l’oxygène. Sa teneur peut atteindre 800 à 2000 ppm selon la plante en C4 : c’est-à-dire des teneurs de 2 à 5 fois plus élevées que sa teneur atmosphérique actuelle.
Dans ces conditions la fixation photosynthétique de l’O2 est faible voire inexistante car les sites actifs de la Rubisco sont tous occupés par le CO2. L’énergie fournie par l’activité des photosystèmes est donc utilisée principalement dans la fixation du CO2 lorsque la température foliaire augmente, expliquant l’optimum thermique plus élevé chez ce type de plante.
Les plantes en C4 ont évolué à partir des plantes en C3 lors de la diminution globale de la teneur en CO2 atmosphérique à la fin de l’ère tertiaire [12].
Cette diminution aurait alors « libérée » la fonction oxygénase de la Rubisco des plantes en C3, entrainant une perte du carbone fixé via la photorespiration.
La mise en place d’un mécanisme de concentration de CO2 est un avantage car elle empêche cette perte de carbone. On trouve actuellement des espèces qui sont des « intermédiaires » entre les C3 et les C4.
5. L’optimum thermique de la photosynthèse C3 est modulé par certains paramètres environnementaux
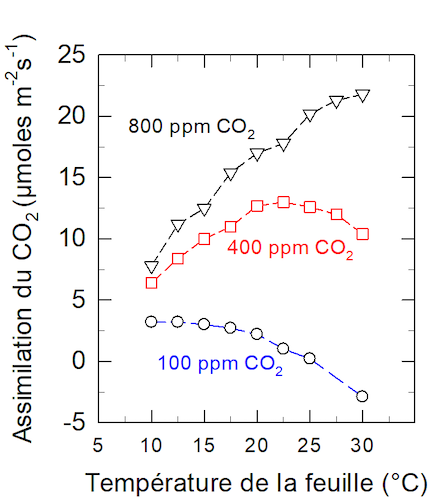
5.1. La teneur en CO2 dans l’atmosphère
L’optimum thermique croit avec l’augmentation de la teneur en CO2 ambiante. Dans le cas présenté sur la Figure 12, elle passe de 10°C environ lorsque sa teneur est de 100 ppm à plus de 30°C lorsqu’elle est de 800 ppm.
L’appareil photosynthétique est résistant à la sécheresse. Il garde toutes ses capacités d’absorption de CO2 sur la Rubisco, et de production d’énergie jusqu’à ce que les feuilles aient perdu environ 30% de leur eau [13].
L’assimilation de CO2 diminue dans cette gamme de perte en eau, parce que les stomates se ferment (Lire Focus Transpiration de la feuille et protection contre la chaleur). Cette fermeture freine l’entrée de CO2dans la feuille et entraine par conséquence une baisse de la teneur en CO2 dans le mésophylle.
Cependant, la teneur en O2 dans les chloroplastes reste toujours élevée. En effet sa teneur dans l’atmosphère (21% soit 210 000 ppm) est, comparativement à celle du CO2 (@ 400 ppm), très élevée et en tout cas suffisante pour qu’une quantité très substantielle traverse l’épiderme même lorsque les stomates sont fermés.
La compétition entre CO2 et O2 pour l’occupation des sites actifs de la Rubisco est donc en faveur de l’O2.
Figure 14. A, Variations de l’assimilation de CO2 en fonction de la température foliaire. Des feuilles ayant perdues des quantités d’eau différentes se trouvant dans un air dont la teneur ambiante en CO2 est de 400 ppm. B, La vitesse de transfert d’électrons estimée sur les mêmes feuilles par la mesure de l’émission de fluorescence chlorophyllienne. [Source : Schéma de l’auteur, d’après Cornic et al. ref. 14]Par conséquent, l’optimum thermique de la photosynthèse doit s’abaisser chez les plantes en C3 qui se dessèchent.
C’est ce que montre la Figure 14A, dans laquelle l’optimum thermique passe de 23°C environ, chez une feuille de Pois au maximum de sa turgescence, à 17°C lorsqu’elle a perdu 20% de son eau.
Le transfert d’électrons dans la membrane des thylacoïdes n’est pas modifié par la perte en eau dans la gamme indiquée (Figure 14B). Lorsque la perte en eau est de 20%, l’énergie produite par l’activité des photosystèmes est principalement utilisée pour fixer l’oxygène atmosphérique sur le RuBP [14], entrainant une augmentation de la photorespiration.
6. Pourquoi, à partir de son optimum thermique, l’assimilation de CO2 diminue lorsque la température s’abaisse ou augmente ?
6.1. Lorsque la température s’abaisse
Plusieurs raisons contribuent probablement toutes, à des degrés divers, à cette diminution :
La vitesse de renouvellement du RuBP diminue : il y a un ralentissement de l’activité de certaines enzymes contrôlant ce renouvellement, notamment celle d’une Fructose 1,6-bisphosphate (voir Figures 9 et 11).
Séquestration des composées phosphorylés dans les chloroplastes. Les trioses phosphate ne sont plus (ou moins) exportés lorsque la synthèse de saccharose est inhibée. Le phosphate inorganique du chloroplaste n’est plus renouvelé entrainant une baisse de la synthèse d’ATP.
Ralentissement de la chaîne de transfert d’électrons (voir section 4.1), dont le résultat est la réduction de la production d’énergie (pouvoir réducteur et ATP).
Chez les plantes en C4 c’est l’activité de la Rubisco qui parait prépondérante, bien que la sensibilité au froid des enzymes participant à l’accumulation de CO2 au niveau de la Rubisco soit bien connue.
6.2. Lorsque la température augmente
Chez les plantes en C3 l’augmentation de la photorespiration diminue la fraction des électrons produits par le PSII et utilisée pour assimiler le CO2. Cependant d’autres facteurs sont en jeu puisque l’assimilation de CO2 mesurée (1) dans une atmosphère où la photorespiration est faible voire nulle (teneur ambiante en O2 de 1%), et (2) mesurée dans une atmosphère normale chez une plante en C4 diminue dans ces deux cas (Figure 10).
Plusieurs raisons peuvent être invoquées :
Le ralentissement de l’activité PSII entrainant celle de la chaîne de transfert d’électrons du PSII au PSI.
Pour assurer son rôle la Rubisco doit être activée par une enzyme appelée Rubisco activase, dont l’activité diminue lorsque la température est supérieure à 33°C environ (d’ailleurs, des activases résistantes aux fortes températures apparaissent chez certaines plantes soumises à des périodes de fortes chaleurs [15]). Cependant comme l’activase doit elle-même être activée par un processus dépendant du transfert d’électrons, il n’est pas exclu que ce dernier participe aussi à la limitation [15].
Les « ratés de catalyse » de la Rubisco augmentent avec la température et des quantités de plus en plus importantes d’un inhibiteur de l’enzyme (le Xylulose-1,4-bisphosphate), dont la structure est proche de celle du RuBP (voir Figures 9 et 11) sont synthétisées.
Chez les plantes en C4 (cas du Maïs) l’activation et l’activité des enzymes qui participent au système de concentration du CO2 au niveau de la Rubisco sont peu sensibles aux températures élevées. Les mêmes raisons que précédemment peuvent expliquer la diminution de l’assimilation du CO2 lorsque la température augmente au-delà de celle de l’optimum thermique.
7. Endurcissement après exposition de la plante aux températures fraiches (≤ 10°C environ) et élevées (≥ 37°C environ)
Le maintien de plantes à des températures fraiches ou élevées provoque, avec les modifications de la photosynthèse décrites précédemment, un accroissement de leur résistance à des températures autrement létales (gel et température élevées). C’est l’endurcissement.
Dans ce processus, température et lumière sont en interaction et les changements métaboliques induits sont parfois très rapides (de la minute à l’heure).
Ainsi, un endurcissement au froid peut être obtenu à température ordinaire en modulant la longueur de la période lumineuse ou sa composition spectrale dans le rouge[16]. Cependant, le froid reste nécessaire pour obtenir un endurcissement complet. De même l’absence de lumière au froid empêche l’endurcissement à des degrés divers.
Aux températures élevées: les signaux transmis activent la synthèse de protéines chaperones (HSPs : Heat Schock Proteins) qui réparent les protéines qui se dénaturent, empêchent aussi leur coagulation ou encore contribuent à les marquer pour dégradation.
Aux températures fraiches: la synthèse des protéines chaperones est aussi activée. Elle est accompagnée (i) de la synthèse de protéines « antigel » qui interfèrent avec la formation des cristaux de glace et (ii) d’une augmentation de la synthèse de sucre tendant à augmenter la pression osmotique dans les cellules.
Noter que les voies de signalisation et leurs interactions induisant la réponse du génome ne sont connus que partiellement. Les références données dans « En savoir plus » et un Focus attaché permettent un approfondissement de ce point en évolution.
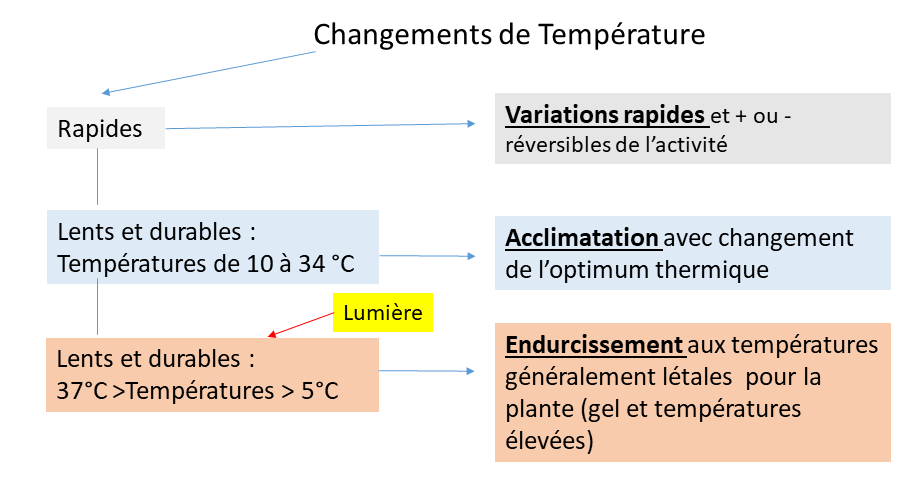
8. Effets de la température sur la photosynthèse : schéma récapitulatif
Le schéma récapitulatif (Figure 15) classe les effets de la température sur la photosynthèse en fonction de la rapidité des changements de la température et de l’étendue de ses variations. A noter que l’endurcissement permet le maintien des feuilles chez les plantes à feuilles pérennes et par conséquent minimise les pertes énergétiques dans des conditions de températures extrêmes.
Figure 15. Schéma classant les effets de la température sur la photosynthèse. [Source : G. Cornic]La rapidité des changements climatiques actuels rend nécessaire l’approfondissement des réactions des plantes à leur environnement : l’espoir est de pouvoir conserver une production primaire suffisante pour le maintien du fonctionnement de la biosphère.
9. Messages à retenir
L’assimilation du CO2 par une feuille présente un optimum thermique proche de la température moyenne de son milieu de croissance.
Cet optimum thermique peut changer rapidement lorsque les conditions du milieu sont modifiées durablement : c’est un processus d’acclimatation.
Cet optimum thermique est en moyenne moins élevé chez les plantes en C3 que chez les plantes en C4 : ceci est principalement dû à la fixation photosynthétique de l’O2 atmosphérique via l’activité de la Rubisco chez les plantes en C3.
Cet optimum dépend de la teneur en CO2 de l’air ambiant chez les plantes en C3: à teneur élevée il devient identique à celui des plantes en C4.
Cet optimum dépend de l’état d’hydratation de la feuille.
Soumises à des températures fraiches ou chaudes les plantes mettent en jeux des processusd’endurcissement aux températures autrement létales. Ces processus impliquent des synthèses de protéines et des modifications de la fluidité des membranes chloroplastiques et cellulaires.
Notes et références
Image de couverture. Coucher de soleil sur le désert de Sonora en Arizona. [Source : libre de droits / Pixabay]
[1] Meehl GA, Stocker TF, Collins WD, Riedlingstein P, Gaye AT, Gregory JM, Kitoh A, Knutti R, Murphy JM, Noda A & Raper SCB (2007). Climate Change 2007: The Physical Science Basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press
[2] Yamori W, Hikosaka K & Way DA. (2014). Temperature response of photosynthesis in C3, C4, and CAM plants: temperature acclimation and temperature adaptation. Photosynthesis Res. 119, 101– 117.
[3] Par exemple, lorsque des plantes en croissance sont soumises à une sécheresse, la quantité de carbone qu’elles assimilent diminue d’abord parce que la croissance foliaire est inhibée. Les mécanismes permettant la fixation de CO2 dans la feuille ne sont pas alors inhibés. Boyer JS (1970) Plant Physiol. 46, 233-235
[4] Les valeurs des optimums thermiques donnés ici, sont issues de mesures faites dans « air normal », contenant 21% d’O2 et environ 400 ppm de CO2. Lorsque cela n’est pas le cas les teneurs en O2 et CO2 sont indiquées. L’assimilation de CO2 dans un air contenant 21% d’O2 est saturée à partir de 1200 ppm de CO2 environ lorsque la lumière est proche de la saturation. Le pouvoir évaporant de l’air est aussi régulé dans la plupart des cas durant les mesures. Il est estimé par le déficit de saturation de la pression partielle en vapeur d’eau dans l’air ambient autour des feuilles.
[5] Plantes issues d’un même individu par reproduction végétative. Ils sont génétiquement identiques.
[6] Ecotype : Plantes d’une même espèce issues de milieux différents, et qui, cultivées de la graine à la fleur dans des conditions identiques présentent des caractéristiques physiologiques différentes.
[7] Elle va chercher l’eau jusqu’à la nappe phréatique d’où sa dénomination de plante phréatophyte.
[8] Pearcy RW (1971). Acclimation of photosynthetic and respiratory CO2 exchange to growth temperature in Atriplex lentiJormis (Torr.) Wats. Plant Physiol. 59, 795-799
[9] Yamasaki T, Yamakawa T, Yamane Y, koike H, Satoh K & Katoh S. (2002) Temperature acclimation of photosynthesis and related changes in photosystem II electron transport in winter wheat. Plant Physiol. 128 1087-1097.
[11] Jordan DB & Ogren WL (1984). The CO2/O2 specificity of ribulose 1,5-bisphosphate carboxylase/oxygenase. Dependence on ribulose bisphosphate concentration, pH and temperature. Planta161, 308-313
[12] Ehleringer JR, Sage RF, Flanagan LB & Pearcy RW (1991). Climate change and the evolution of C4 photosynthesis. Trends in Ecology and Evolution 6, 95-99
[13] Cornic G & Massacci A (1996). Leaf photosynthesis under drought stress. In Advances in Photosynthesis (vol 5) Photosynthesis and the environment, 347-366. Neil R Baker (ed.) Kluwer Academic publishers Dordrecht.
[14] Cornic G, Badeck F-W, Ghashghaie J & Manuel N (1999). Effect of temperature on net CO2 uptake, stomatal conductance for CO2 and quantum yield of photosystem II photochemistry of dehydrated pea leaves. In Sanchez Dias M, Irigoyen JJ, Aguirreolea J & Pithan K (eds) Crop development for cool and wet regions of Europe. European community. ISBN 92-828-6947-4.
[15] Crafts-Brandner SJ, van de Loo FJ & Salvucci ME (1997). The two forms of ribulose-1,5-bisphosphate carboxylase/oxygenase activase differ in sensitivity to elevated temperature. Plant Physiol. 114, 439-444.
[16] Puhakainen T, Li C, Boije-Malm M, Kangasjärvi J, Heino P & Palva ET. (2004). Short-day potentiation of low temperature-induced gene expression of a C-repeat-binding factor-controlled gene during cold acclimation in Silver Birch. Plant Physiol. 136, 4299-4307
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.
Le changement climatique global est un sujet de préoccupation majeur. Il part du constat fait…
Serge PLANTON, Ingénieur général des ponts, des eaux et des forêts, ancien chercheur climatologue à Météo-France, CNRM (Centre National de Recherches Météorologiques).