



Le génome entre stabilité et variabilité
PDF
En permanence, le génome des êtres vivants est affecté par des anomalies, sous l’effet d’erreurs fortuites ou d’agents internes et externes à l’organisme. Heureusement, des enzymes corrigent la plupart de ces anomalies et confortent donc la stabilité génétique. Mais elles peuvent aussi être à l’origine de mutations qui participent de la variabilité. D’autres systèmes permettent, eux, de produire de nouvelles combinaisons génétiques. Ces mutations et recombinaisons sont indispensables pour l’adaptation des populations aux modifications du milieu et donc pour l’évolution.
1. Introduction
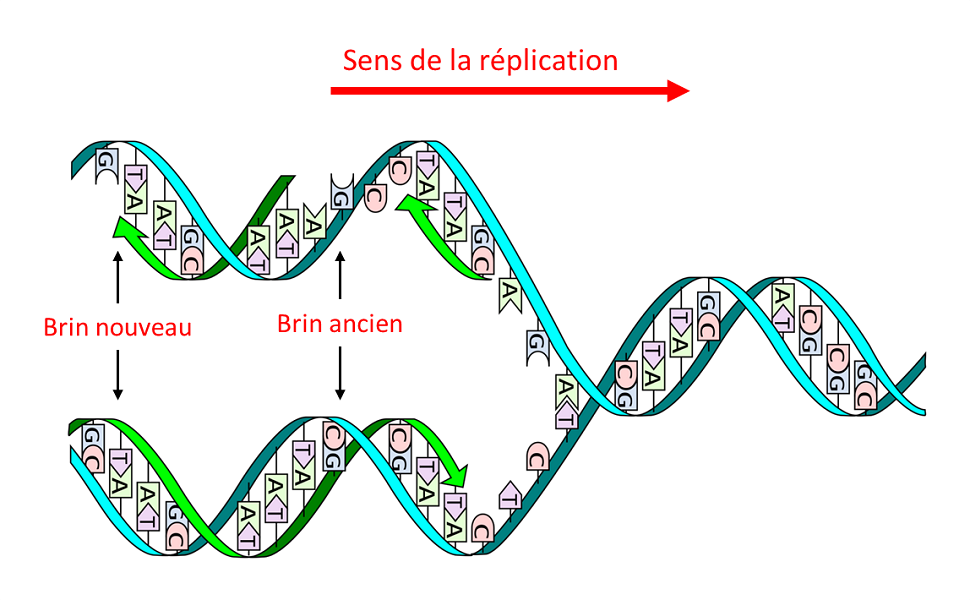
L’ADN est constitué d’un double enchaînement de monomères (les nucléotides) dont il existe quatre types qui ne diffèrent que par la base azotée qu’ils portent : adénine (A), guanine (G), thymine (T) et cytosine (C). C’est la séquence précise de ces quatre types, et donc des quatre bases, qui constitue le « texte » du message génétique. Sa structure en deux brins complémentaires, où A est toujours face à T et G face à C, – la fameuse double hélice (Figure 1) – présente deux avantages majeurs :
- Elle permet, en une seule étape, la réplication en deux molécules filles identiques (Figure 1), donc la transmission de l’information génétique à travers les divisions cellulaires et la reproduction sexuée.
- Elle permet la réparation des lésions apparues sur l’un des brins ainsi que la création de molécules recombinantes, c’est à dire de nouvelles combinaisons génétiques. Cette structure double-brin favorise donc à la fois la stabilité et la variabilité des génomes.
Les variations génétiques de l’ADN se rangent en deux grandes catégories : les mutations et les recombinaisons.
- Les mutations sont des changements brusques dans le génome d’une cellule vivante ou d’un virus. Chez les organismes pluricellulaires, si les mutations touchent les cellules germinales destinées à donner les gamètes, elles seront transmises à la descendance et deviendront donc héréditaires.
- Les recombinaisons consistent à produire une nouvelle combinaison génétique à partir d’échanges entre matériels génétiques existants. Il en existe plusieurs types dont les mécanismes et les rôles biologiques sont très différents. Classiquement, cette définition ne concernait que des ADN appartenant à la même espèce. On sait maintenant qu’il existe des échanges génétiques spontanés entre espèces différentes, la transgenèse, que nous inclurons dans cette catégorie.
Ces deux grands types de variations se divisent eux-mêmes en plusieurs modalités. Nous nous efforcerons d’en extraire les éléments les plus à même d’éclairer leurs rôles respectifs dans la dynamique des populations et l’évolution.
2. Les mutations
2.1. Généralités
La stabilité des messages génétiques codés par l’ADN peut être affectée par des erreurs de l’enzyme qui effectue la réplication, la réplicase. Cette enzyme peut ainsi mettre une base azotée à la place d’une autre sur le brin nouveau. Mais en dehors de toute réplication, l’ADN subit aussi en permanence des lésions. Il faut bien faire la différence entre lésion et mutation (lire Polymorphisme génétique & variation). Les lésions sont des anomalies de la structure physique de l’ADN qui, la plupart du temps, vont empêcher sa réplication. Elles ne peuvent donc être transmises à la descendance. Par contre, une molécule mutée a une structure physique normale. Seule la séquence des bases – et donc l’information qu’elle porte – est modifiée. Elle peut donc se répliquer sans problème et transmettre la mutation aux cellules filles.
Les lésions sont très diverses et il existe des quantités d’enzymes de réparation, chacune étant très spécialisée vis-à-vis d’un type particulier de lésion. On parle à ce propos de « boîte à outils des cellules » pour permettre la réparation de l’ADN (travaux récompensés par le prix Nobel de Chimie 2015). Assez paradoxalement, ce sont ces systèmes de réparation qui, par erreur, créeront la mutation proprement dite à partir de la lésion primaire. Ces lésions peuvent être provoquées soit par des agents internes à l’organisme (ou endogènes) soit par des agents du milieu extérieur (ou exogènes). Voici quelques exemples pour fixer les idées.
- Agents endogènes : chez les animaux à sang chaud, on estime que la molécule d’ADN peut subir de 20 000 à 40 000 coupures simple-brin par cellule et par jour, sous l’effet de l’agitation moléculaire. Il peut y avoir aussi des pertes de bases, on estime à 10 000 le nombre des T et C perdues et à 500 celui des A et G, toujours par cellule et par jour. Dans ces cas-là on peut même parler de lésions spontanées. Parmi les agents endogènes les plus fréquents, citons des dérivés de l’oxygène (radicaux libres ou ROS), qui sont des sous-produits normaux du métabolisme respiratoire, ils jouent un rôle important en oxydant certaines bases qui devront donc être remplacées. Il faudrait aussi rajouter les éléments transposables, nous en reparlerons plus loin.
- Agents exogènes : ils peuvent être physiques (radiations) ou chimiques. Les plus courants sont les rayons ultra-violets (UV), dont l’effet est généralement limité à la peau car ils sont peu pénétrants (lire Impact cellulaire des UV solaires). A titre d’exemple, on estime qu’un bain de soleil peut provoquer, par cellule et par heure, de 60 000 à 80 000 liaisons chimiques anormales entre des thymines contiguës d’un même brin d’ADN, dont chacune suffit à bloquer la réplication. Si elles ne sont pas toutes réparées, la cellule meurt : c’est le coup de soleil.
On voit donc bien que la stabilité de l’ADN est un processus dynamique, résultat d’un équilibre permanent entre la production de lésions et leur réparation. Ces mécanismes de réparation ne fonctionnent pas à niveau constant, ils sont soumis à régulation.
Un premier type de régulation dépend du nombre de lésions dans la cellule. Ce phénomène a d’abord été mis en évidence chez le colibacille, bactérie qui est un des objets d’étude préféré des généticiens. Dés 1974, avait été postulée l’existence d’une réponse, dite SOS [2], qui régule l’intervention de plusieurs systèmes de réparations en fonction du nombre de lésions. Lorsqu’il y a un faible nombre de lésions, cette réponse augmente l’efficacité des mécanismes de réparations fidèles. Mais au-delà d’un certain seuil de lésions, ces mécanismes sont débordés. La réponse SOS induit alors la synthèse d’une réplicase capable de franchir des lésions ponctuelles (voir plus loin), mais avec un certain taux d’erreurs. C’est ce que l’on dénomme la mutagenèse SOS. A la fin des années 1990, il a été démontré qu’elle facilite l’adaptation de la population bactérienne à un milieu devenu hostile, au prix bien sûr de pertes importantes dues aux mutations délétères. C’est en quelque sorte un système de la dernière chance, d’où l’appellation de SOS.

Une induction des mécanismes de réparations fidèles suite à des irradiations a été mis en évidence sur d’autres organismes très prisés des biologistes (Figure 2) : dans les années 80 chez la levure de boulangerie, en 2000 chez la mouche du vinaigre [3], en 2006 chez une plante au joli nom, « l’arabette des dames » [4] et en 2008 chez des souris exposées aux radiations à Tchernobyl [5]. Ces données remettent en cause l’affirmation encore très répandue que les effets des radiations sont directement proportionnels aux doses. Pour des doses modérées, c’est manifestement faux. Sur ces travaux, on trouvera des informations plus détaillées dans : https://www.lespiedsdansleplat.me/comment-les-organismes-vivants-protegent-leur-adn/.
Le second type de régulation porte sur la fidélité de la réplication selon le type d’organisme. Dans toutes les cellules vivantes, bactéries et autres, cette réplication fait intervenir, en plus de la réplicase-ADN, un système enzymatique correcteur. Grâce au brin ancien, ce système corrige les erreurs commises sur le brin nouveau par la réplicase ce qui aboutit à un taux d’erreur très faible, de l’ordre de 1 sur 10 milliards (10-10). Une telle précision est indispensable pour des organismes ayant de grandes molécules d’ADN : 4 millions de paires de bases chez le colibacille et 3 milliards dans l’espèce humaine.
Mais chez les organismes pluricellulaires, le nombre de cellules peut être très grand : dans l’espèce humaine, il est estimé à cent mille milliards (1014). Chez ces organismes, la majorité des mutations sont neutres, car le génome comprend une grande quantité de séquences non-codantes (lire Polymorphisme génétique et variation). Même en tenant compte de cela, un taux d’erreur de 1 sur 10 milliards (10-10) est encore trop important. Et ceci d’autant plus que de nombreuses mutations se produisent aussi hors réplication. S’il n’y avait qu’un seul exemplaire de gène par cellule, beaucoup d’entre elles porteraient des mutations délétères et l’organisme ne serait pas viable. D’où l’intérêt, et même la nécessité, d’avoir deux lots chromosomiques pour ces organismes (diploïdie). La plupart des mutations qui inactivent un gène étant récessives, il suffit que l’autre exemplaire soit fonctionnel pour que la cellule fonctionne normalement. En simplifiant, on peut dire que la diploïdie joue le rôle d’une roue de secours.
S’il l’on s’intéresse maintenant au cas des virus, dont la plupart ont de très petits génomes, la situation est très différente. Pour les virus à ADN, il n’y a généralement pas d’activité correctrice associée à la réplicase, le taux de mutation est donc beaucoup plus élevé, de 10-6 à 10-8 selon les virus. Ceci est encore plus vrai pour les virus dont le génome est ARN, car les réplicases ARN sont beaucoup moins fidèles que les réplicases ADN, le taux d’erreur est alors de 10-6 à 10-4. Cependant, chez ces virus ARN, il faut distinguer deux catégories en fonction de la taille du génome. Chez ceux dont la taille du génome se situe autour de 10 000 nucléotides, la fidélité de la réplication dépend de la seule réplicase. C’est le cas des virus de la grippe (13 500 nucléotides) et du VIH, responsable du SIDA (10 000 nucléotides). Chez ce dernier, le taux de mutations ponctuelles a été estimé à 1 sur 10 par génome et par cycle de réplication. Comme le nombre de particules virales produites (virions), est de l’ordre de 10 000 par jour et par cellule infectée, on voit que la population globale chez un hôte infecté peut présenter une variabilité considérable.
Chez les virus ARN dont le génome situe autour de 30 000 nucléotides ou plus, on sait maintenant qu’il y a une activité correctrice associée à la réplicase. C’est le cas des Coronavirus, dont le SARS-CoV-2 responsable de la pandémie de COVID-19, dont le génome contient 29 903 nucléotides. Cette activité correctrice améliore d‘un facteur 20 la fidélité de réplication, permettant ainsi de ne pas produire un nombre excessif de virions défectifs.
Pour dire les choses de façon plus générale, chez tous les virus, qu’ils soient ADN ou ARN, il semble bien y avoir une corrélation négative entre le taux de mutations et la taille du génome. Pour chaque espèce virale s’est élaboré, par sélection naturelle, un équilibre pour le taux de mutations. Il ne doit pas être trop élevé, pour que la perpétuation de l’espèce virale ne soit pas compromise, mais assez quand même pour créer une variabilité qui permet au virus de contourner les mécanismes de défense de leur hôte. Sur ce dernier point le cas du VIH est tout à fait emblématique.
Ceci nous amène à faire deux remarques sur le rôle des mutations dans le processus évolutif :
- Elles jouent un rôle clé dans l’adaptation des espèces à leur milieu car, en augmentant la diversité génétique, elles sont le matériau sur lequel va pouvoir agir la sélection naturelle. Ce que nous venons de voir avec les virus en est un exemple. Le développement des bactéries résistantes aux antibiotiques, des insectes aux insecticides, ou des plantes aux herbicides, en sont d’autres.
- Une mutation génétique n’est pas avantageuse ou désavantageuse en soi, tout dépend de l’environnement. A titre d’exemple, aux îles Kerguelen, dans le sub-antarctique, on trouve une espèce de mouche sans ailes, Calycopteryx moseley (Figure 3). Ce caractère, fortement désavantageux dans nos contrées, est au contraire bénéfique là-bas car il évite à ces mouches d’être emportées dans l’océan par les vents très forts qui balayent ces îles en permanence.

Figure 3. Mouche sans ailes, Calycopteryx moseley, vivant aux Iles Kerguelen, dans un milieu extrêmement venté. [source : Photos © Marine Pouvreau].

Quelles sont les différentes catégories de mutations ? On les distingue classiquement par la taille du segment d’ADN concerné. Par ordre croissant, cela va des mutations ponctuelles, où une seule paire de bases azotées est modifiée, aux mutations qui touchent des segments d’ADN plus ou moins grands, jusqu’à plusieurs dizaines de milliers de nucléotides. Sans oublier les modifications du nombre de chromosomes. Les deux dernières catégories concernent principalement les organismes eucaryotes, dont les chromosomes sont localisés dans un compartiment particulier de la cellule : le noyau. Ces chromosomes sont chacun constitués d’une très longue molécule d’ADN mais dans un « emballage » complexe, composé de plusieurs familles de protéines.
2.2. Les mutations ponctuelles
Elles peuvent être dues soit à des erreurs lors de la réplication soit, plus fréquemment, à des lésions mal réparées ; elles créent des variants d’un même gène (allèles). Chez le colibacille, c’est le système SOS qui provoque la grande majorité des mutations après irradiation aux UV. La plupart des exemples cités plus haut relèvent en fait de mutations ponctuelles.
2.3. Les mutations chromosomiques
Ce sont des réarrangements produits par des agents qui provoquent des cassures sur l’ADN, dont les radiations (autres que les UV). Plusieurs cassures double-brin peuvent aboutir à des réarrangements plus ou moins importants selon leur nombre et la taille du segment concerné.
Dans le cas de deux cassures double-brin très éloignées sur le même chromosome, c’est tout le segment situé entre les cassures qui pourra être soit (a) perdu, entraînant la létalité de l’individu, (b) être inversé au même site (inversion), ou (c) transféré sur un autre chromosome (translocation) si d’autres cassures se sont produites sur celui-ci. Inversions et translocations sont assez fréquentes dans les populations naturelles. Elles gênent le bon appariement des chromosomes lors de la méiose et entraînent donc une certaine stérilité. De ce fait, elles peuvent intervenir dans les processus de spéciation. Souvent, des espèces voisines diffèrent par des réarrangements chromosomiques.
2.4. Les mutations du lot chromosomique
Cette catégorie obéit à des mécanismes différents des précédentes. Il ne s’agit plus d’anomalies résultant de lésions primaires mais d’erreurs mécaniques dans les processus de division cellulaire, mitose ou méiose. Certaines peuvent aboutir à des situations où les individus ont plus de deux lots chromosomiques (2N), mais toujours un nombre entier de lots (3N, 4N…), c’est la polyploïdie (lire Focus Polyploïdie). Ils sont viables car la « balance génique » est respectée : tous les gènes possèdent bien le même nombre d’exemplaires. Par contre, les 3N sont stériles car la balance génique des gamètes est forcément anormale. La méiose ne peut évidemment pas répartir équitablement entre les gamètes un nombre impair de chromosomes. Les polyploïdes 4N, 6N ou 8N quant à eux sont fertiles. Mais ils créent d’emblée une nouvelle espèce, car tout croisement avec les parents diploïdes donnerait des descendants stériles, leurs méioses étant très déséquilibrées.
D’autres « ratés » de la méiose peuvent aboutir à un déséquilibre chromosomique dans certains gamètes, qui se retrouvera donc dans les descendants. Ils seront par exemple 2N-1 (monosomie) ou 2N+1 (trisomie), on parle alors d’aneuploïdie, qui est une situation délétère car la balance génique n’est plus respectée.
3. Les recombinaisons
Il existe de nombreuses modalités de recombinaison dont les rôles biologiques sont très différents.
3.1. La recombinaison homologue
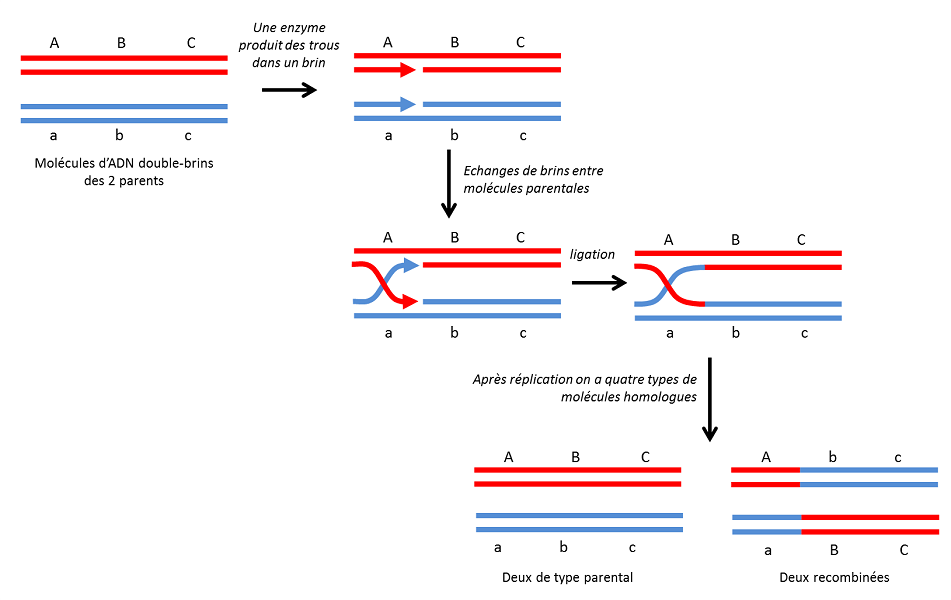
C’est le type le plus courant et aussi le plus anciennement connu. Comme son nom l’indique, elle se fait entre molécules d’ADN identiques (Figure 4). Chez les organismes diploïdes, elle se produit régulièrement, lors de la méiose, entre chromosomes homologues, par enjambement (crossing-over). C’est une variabilité « douce » : le recollement se fait exactement au même point donc ne s’accompagnera ni de mutations, ni de modifications dans l’agencement des gènes sur les chromosomes. On obtient ainsi de nouvelles combinaisons génétiques tout à fait viables qui pourront, dans un environnement donné, avoir des capacités adaptatives différentes de celle des combinaisons parentales. Chez les bactéries, elle peut également se produire lors de la conjugaison [6] qui permet des échanges génétiques entre cellules [7].
Ce mécanisme de recombinaison intervient aussi dans la réparation de l’ADN. Chez les organismes diploïdes, les cassures double-brin provoquées sur un chromosome par une irradiation peuvent être réparées grâce à la présence d’une molécule homologue intacte dans le segment concerné.
3.2. Autres types de recombinaisons
Pour simplifier, nous regrouperons là des types d’événements aux rôles biologiques très différents. On pourrait les réunir sous le terme de recombinaison additive car ils aboutissent, dans la majorité des cas, à des additions de segments d’ADN dans les génomes.
Parmi les mécanismes les plus courants, il faut mentionner la transposition des éléments génétiques mobiles (appelés éléments transposables) [8], qui ne comportent que quelques centaines ou milliers de nucléotides et possèdent, au minimum, des fonctions nécessaires à leurs « sauts » dans le génome hôte. Ils se rencontrent aussi bien chez les bactéries que chez les eucaryotes les plus complexes, parfois en grand nombre. Chez les premières, ils peuvent s’échanger entre espèces différentes lors de la conjugaison [6]. Comme ils portent généralement des gènes de résistance aux antibiotiques, ils sont l’un des facteurs essentiels de la propagation très rapide de ces résistances chez les bactéries pathogènes, avec les graves problèmes médicaux qui en découlent.
Dans l’espèce humaine, alors que nos gènes proprement dits, qui codent pour toutes nos protéines, ne constituent qu’environ 2 à 3% de l’ADN du noyau, les diverses familles d’éléments transposables en constituent près de 50 %. Elles sont pour la plupart installées dans notre filiation depuis des temps géologiques très reculés, bien avant l’apparition de notre espèce. La famille des rétrovirus, à laquelle appartient le VIH, à la fois virus et élément transposable, représente à elle seule 8 % de notre ADN. Fort heureusement pour la stabilité des génomes, un petit nombre seulement de ces éléments sont encore mobiles. Comme pour les mutations « classiques », l’insertion d’un élément transposable peut être délétère si elle se fait dans un gène, mais elle peut aussi apporter des innovations génétiques intéressantes, soit en modifiant des régulations de gènes, soit par leurs propres fonctions. De multiples données montrent qu’ils ont contribué à l’évolution des génomes donc des espèces [9].
Enfin, il faut mentionner les phénomènes de transgenèse, c’est à dire les échanges génétiques entre espèces différentes. Depuis la fin des années 1990, on s’est aperçu qu’ils sont beaucoup plus fréquents qu’on ne le pensait. La barrière d’espèce, que l’on avait crue infranchissable dans les conditions naturelles, est en fait assez poreuse, du moins à l’échelle évolutive. Là encore, on a une source d’innovations qui contribue à la plasticité des génomes et à l’évolution.
4. Conclusion
Nous avons présenté un rapide survol des différents types de variations des génomes. Nous n’avons pu détailler les systèmes enzymatiques impliqués dans les réparations et recombinaisons, ils auraient nécessité à eux seuls un article complet (sinon deux !). Le lecteur intéressé par ces mécanismes pourra se reporter aux sites suivants [10]. Pour conclure, deux points importants sont à souligner :
- Les variations génétiques se produisent de manière aléatoire. Elles ne sont pas dirigées par le milieu à des fins adaptatives, comme cela était postulé dans la vision lamarckienne de l’évolution. Ceci a été abondamment démontré expérimentalement (Lire Lamarck et Darwin : deux visions divergentes du monde vivant). Cependant, contrairement à ce que l’on croyait jusqu’aux années 1980, leurs fréquences peuvent être modulées par l’environnement. Nous en avons vu une illustration chez les bactéries avec l’augmentation du taux de mutations par la réponse SOS et, chez la drosophile et l’arabette, avec la stimulation de la recombinaison homologue suite à des irradiations. Celles-ci ne constituent pas le seul facteur capable de moduler la fréquence des variations, de nombreux stress physiologiques peuvent aboutir à ce résultat. A titre d’exemple, l’application d’antibiotiques sur des cultures bactériennes déclenche aussi la mutagenèse SOS et augmente donc la fréquence des mutations [11,12]. Ce sujet est également abordé dans l’article Adaptation : répondre aux défis de l’environnement.
- Cet article se situe au cœur de ce que l’on peut appeler la dialectique stabilité/variabilité du vivant. Deux propriétés a priori antagonistes mais en fait tout à fait complémentaires. Elles sont toutes deux indispensables pour la survie des populations et l’évolution des espèces.
La stabilité pour permettre l’adaptation durable d’une population à son environnement quand celui-ci est relativement stable et la variabilité pour faciliter les changements génétiques lorsque l’environnement se modifie, permettant à la sélection naturelle d’œuvrer. La réponse SOS chez les bactéries en est une bonne illustration puisqu’elle est capable de remplir l’une ou l’autre de ces deux fonctions, selon les conditions de milieu.
Références et notes
Photo de Couverture : © vitstudio ; Image 134698571 via Shutterstock
[1] http://atlasgeneticsoncology.org/Educ/DNAID30001FS.html
[2] R. Devoret (1993) Mécanisme de la mutagenèse SOS, Med/Sci vol.3, n°9, I-VII.
[3] J. Ducau et al. (2000) Mutation Research 460, 69-80
[4] J. Molinier et al. (2006) Nature 442, 1046-1049
[5] B.E. Rodgers & K.M. Holmes (2008) Dose response 6, 209-221
[6] http://www.perrin33.com/microbiologie/genetique/conjugaison_3.php
[7] Vidéo de Miroslav Radman : https://www.reseau-canope.fr/corpus/video/la-recombinaison-genetique-129.html
[8] D. Anxolabéhère, D. Nouaud & W.J. Miller (2000) Éléments transposables et nouveautés génétiques chez les eucaryotes. Med/Sci, I n° 11, vol. 16, I-IX.
[9] voir « Les surprises de l’évolution : des virus parmi nos ancêtres ? » dans www.lespiedsdansleplat.me
[10] http://www.cours-pharmacie.com/biologie-moleculaire/reparation-de-ladn.html ou http://gec.sdv.univ-paris-diderot.fr/genetique/chapitre9.html
[11] S. Da Re & M.-C. Ploy (2012) Antibiotiques et réponse SOS bactérienne. Med Sci (Paris) 28:179–184.
[12] J. Blázquez, J. Rodrı́guez-Beltrán & I. Matic (2018) Antibiotic-Induced Genetic Variation: How It Arises and How It Can Be Prevented. Annu. Rev. Microbiol.72:209–30.
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Pour citer cet article : BREGLIANO Jean-Claude (30 mai 2021), Le génome entre stabilité et variabilité, Encyclopédie de l’Environnement. Consulté le 26 mars 2025 [en ligne ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/vivant/genome-entre-stabilite-variabilite/.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.


