



Résistance des plantes aux stress : rôle de la respiration ?
PDF
Oui, les plantes respirent ! Leur croissance résulte d’un délicat équilibre entre l’acquisition photosynthétique de carbone et la restitution respiratoire d’une partie de celui-ci. Lors de la photosynthèse, les végétaux absorbent le dioxyde de carbone, CO2, dont la présence croissante dans l’atmosphère pose problème, et rejettent du dioxygène, O2. Cet échange gazeux qui caractérise l’autotrophie des plantes, c’est-à-dire leur capacité à fabriquer des composés organiques à partir d’eau, de minéraux et de CO2 en présence de lumière. Au cours de la respiration, les plantes dégradent une partie des composés formés par la photosynthèse et produisent de l’énergie ; elles consomment alors de l’oxygène et libèrent du CO2. C’est grâce à cela que la cellule végétale réalise la synthèse des nombreux composés constitutifs de sa biomasse. Mais que se passe-t-il en cas de stress ? Comment les mécanismes respiratoires permettent-ils la synthèse de composés de défense, et contribuent-ils alors à une réponse adaptée des plantes à ces stress ?
1. Les échanges gazeux des plantes
La photosynthèse d’une plante ne concerne que les parties chlorophylliennes aériennes, c’est-à-dire principalement les feuilles et les tiges vertes, lorsque la plante est jeune.
Au contraire, la respiration concerne toute la plante. C’est l’équivalent d’une réaction de combustion -elle utilise du dioxygène et libère du dioxyde de carbone- qui permet de transformer les composés carbonés comme le glucose en énergie. Les parties souterraines, donc les racines, respirent aussi. Il existe en effet une atmosphère du sol qui, en milieu bien drainé, contient environ 19% d’O2 (21% dans l’atmosphère), 79% d’azote N2 (78% dans l’atmosphère) et 0,3% de CO2 (0,04% dans l’atmosphère), conditions favorables au processus respiratoire. La respiration des parties aériennes, concerne feuilles et tiges, sans oublier fruits et graines.
Pour une plante annuelle, les échanges gazeux se répartissent donc entre :
- La photosynthèse, qui a lieu exclusivement le jour dans les parties aériennes. Elle est dominante mais pas exclusive puisqu’une respiration aérienne co-existe ;
- La respiration souterraine, essentielle puisque son rôle est de dégrader les sucres (saccharose essentiellement) transmis via la sève élaborée depuis les feuilles jusqu’aux racines.
Cela n’obère pas l’intérêt de la respiration des feuilles, qui contribue à leur vie cellulaire et tissulaire (voir paragraphe 2). La respiration a lieu nuit et jour, contrairement à la photosynthèse.
Tableau 1. Échanges gazeux des différents organes d’une plante. PB : Photosynthèse brute ; RJ : Respiration de jour ; RN : Respiration de nuit, ; PR: Photorespiration.
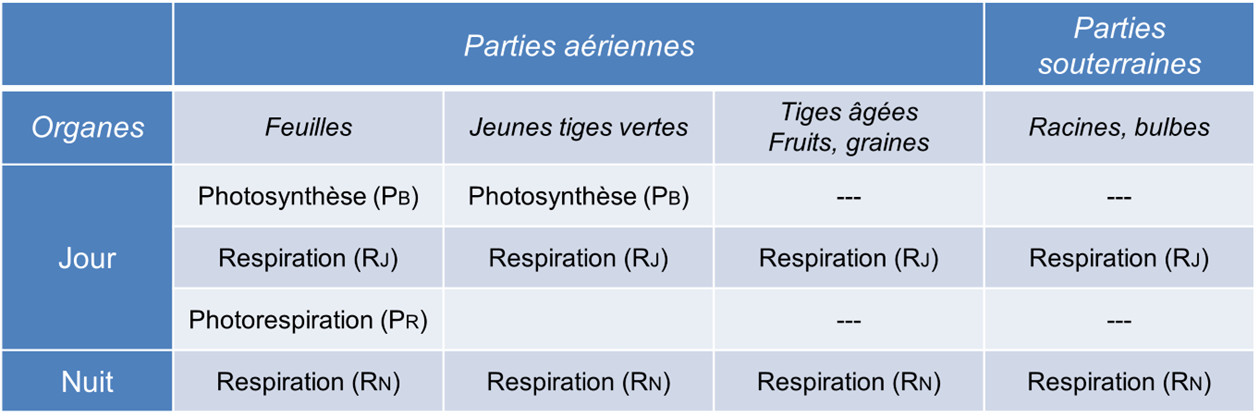

- La photosynthèse (PB) n’a lieu que le jour à la lumière et mobilise exclusivement les chloroplastes.
- La respiration a lieu jour et nuit et mobilise les processus dégradatifs du cytosol (glycolyse, voie des pentoses phosphates) et les mitochondries. Les respirations de jour (RJ) et de nuit (RN) peuvent cependant montrer des intensités différentes (voir paragraphe 2.1).
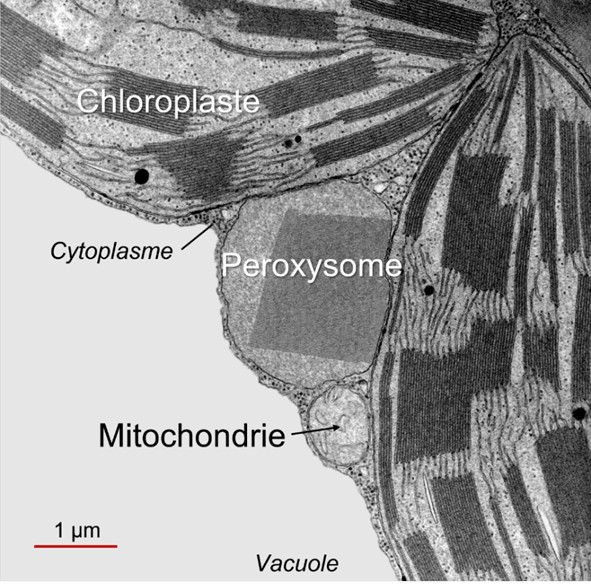
- La photorespiration (PR) ne se déroule qu’à la lumière et fait intervenir les chloroplastes, les peroxysomes et les mitochondries (Figure 1, Lire Le chemin du carbone dans la photosynthèse) (voir paragraphe 2.2). La photorespiration fonctionnant au mieux à 21% d’O2, il est vraisemblable qu’elle soit très faible dans les tiges vertes qui contiennent un ratio O2/CO2 inférieur à celui de l’atmosphère.
La Figure 2 montre un exemple de mesure des échanges gazeux (absorption ou émission de CO2) de jeunes plants de vigne [1] :
- Pendant la journée (période diurne), la courbe représente la photosynthèse nette (PN) ; les processus photosynthétiques sont prépondérants par rapport aux processus respiratoires.
- Pendant la période obscure, seule la respiration fonctionne. La respiration de nuit (RN) représente alors à peine 20% de la photosynthèse nette (PN), et un pourcentage encore plus réduit par rapport à la photosynthèse brute (PB), si l’on tient compte des phénomènes respiratoires qui se déroulent le jour (respiration de jour RJ et photorespiration PR), dont nous reparlerons plus loin (voir paragraphe 2).
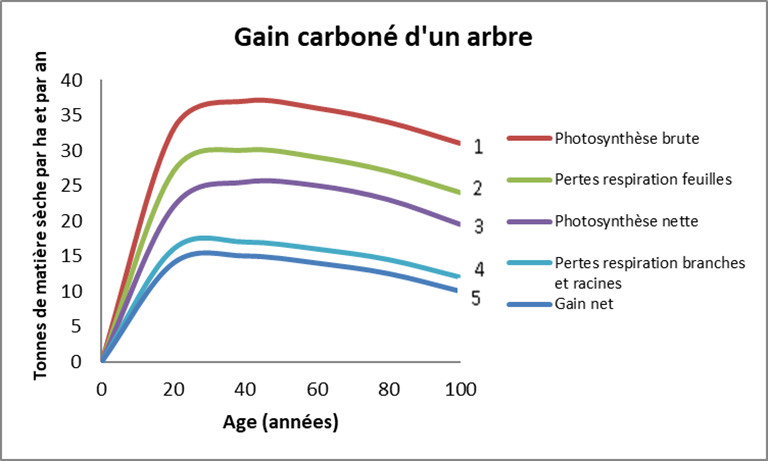
- La courbe 1 représente la photosynthèse brute (PB) des parties aériennes.
- La prise en compte de la respiration (RJ) des feuilles donne la courbe 2, plus basse que la courbe 1.
- La soustraction supplémentaire des pertes de feuilles aboutit à la photosynthèse nette (PN) de l’arbre en courbe 3.
- La courbe 4 prend en compte la respiration du tronc, des branches et des racines.
- Enfin, le gain net (courbe 5) est obtenu en soustrayant les pertes de branches et racines : c’est la courbe la plus basse qui traduit le bilan total.
La figure 2 montre que le gain de carbone par un arbre comme le hêtre est très important dans sa jeunesse (jusqu’à 20 ans), pour atteindre ensuite un plateau [2]. Ce mode de fonctionnement peut être altéré par les contraintes environnementales.
2. Place de la respiration dans les échanges gazeux foliaires
2.1. Bilan des échanges gazeux
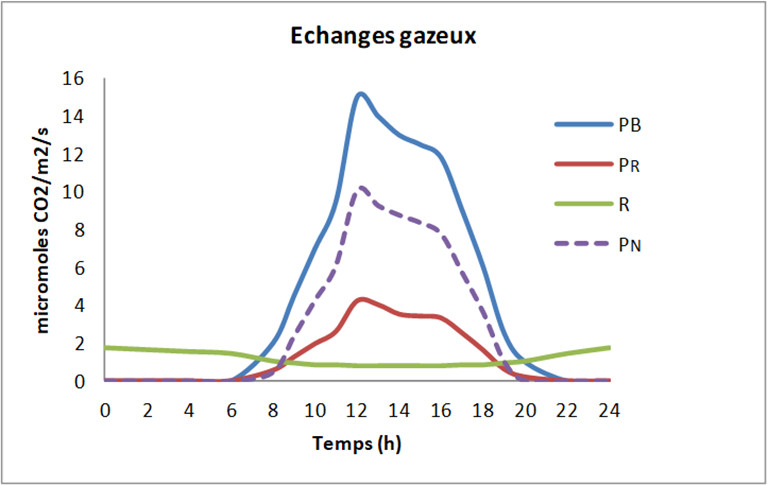
La figure 4 résume les différents échanges gazeux se déroulant dans des feuilles au cours d’un cycle de 24 heures :
- Le processus photosynthétique chloroplastique (photosynthèse brute PB) a lieu le jour et assimile le CO2 tout en rejetant de l’O2.
- La respiration (R), au contraire, consomme de l’O2 et rejette du CO2, et, toujours contrairement à la photosynthèse, a lieu nuit et jour. La respiration est souvent dénommée respiration mitochondriale car les mitochondries sont au cœur de ce processus. Il est aussi important de noter que la respiration de jour (RJ) est plus faible que la respiration de nuit (RN).
- Enfin, un processus assez particulier se rajoute aux deux précédents : la photorespiration (PR) qui ne se déroule que le jour, consomme de l’O2 et rejette du CO2.
La photorespiration représente environ 20 à 25% de la photosynthèse brute chez les plantes de type C3 (Lire Le chemin du carbone dans la photosynthèse). En ce qui concerne la respiration, l’intensité respiratoire nocturne (RN) atteint 10 à 15% de la photosynthèse brute.
La photosynthèse nette (PN), mesurée pendant la phase diurne, est représentée par la formule :
- PN = PB – (PR + RJ)
et pourrait être écrite :
- PN = PB – (0,25 PB + 0,05 PB) soit PN = PB – 0,3 PB
ce qui signifie que les processus de type « respiratoire », provoquant absorption de O2 et rejet de CO2, amputent la photosynthèse de 30% de son rendement.
2.2. La photorespiration ou cycle photorespiratoire
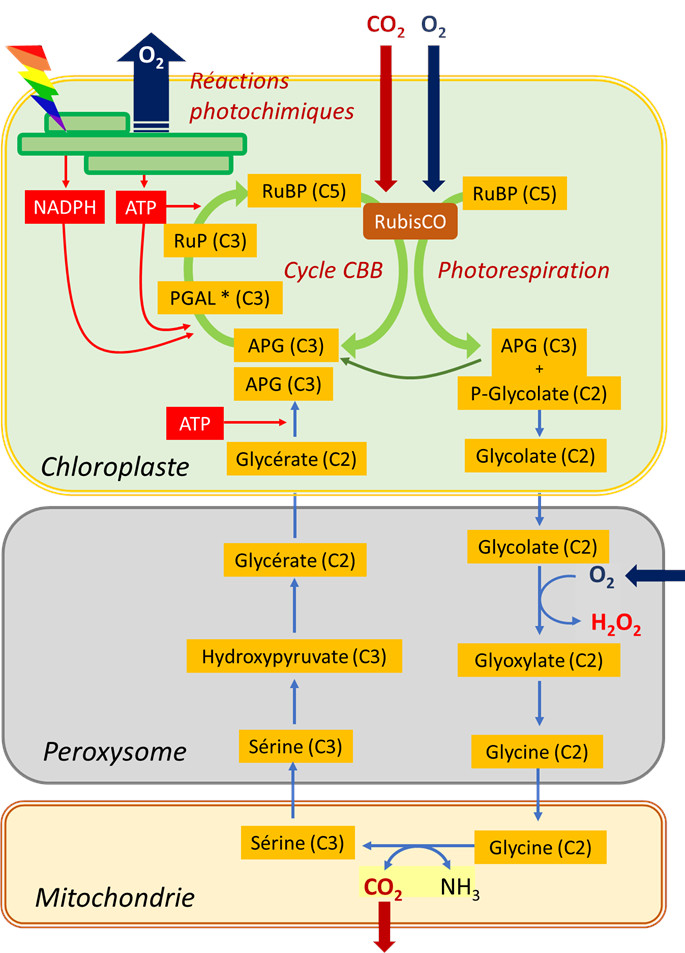
- Par sa fonction carboxylase, la RubisCO intègre une molécule de CO2 sur une molécule de ribulose bisphosphate (molécule en C5) et produit des composés carbonés à 3 atomes de carbone (C3) : acides phosphoglycériques (APG) puis des trioses phosphates (PGAL).
- Par sa fonction oxygénase, la RubisCO intègre une molécule d’O2 à la molécule de ribulose bisphosphate (molécule en C5) qui se coupe en APG (molécule en C3) et phosphoglycolate (composé carboné à deux atomes de carbone, C2). L’APG peut rejoindre le pool des APG issus de la fonction carboxylase tandis que la formation de phosphoglycolate marque le début du cycle photorespiratoire.
Ce cycle voit (i) une succession de composés C2 sortir du chloroplaste et (ii) traverser le peroxysome avant d’être (iii) transformés en un acide aminé en C2, la glycine, dans la mitochondrie. C’est (iv) dans la mitochondrie que du CO2 sera rejeté (ainsi que de l’ammoniac NH3) lors d’une réaction enzymatique complexe qui donnera de la sérine, acide aminé en C3. Le (v) retour vers le chloroplaste (d’où la dénomination de cycle) s’effectue alors en (vi) retraversant le peroxysome, donnant finalement (vii) un APG qui peut encore grossir le pool de ces composés au sein du chloroplaste.
Cette succession de réactions biochimiques :
- est globalement consommatrice d’énergie (ATP) ;
- n’assimile pas de CO2 mais au contraire aboutit à une perte nette de carbone par émission de CO2.
Il faut aussi noter une consommation additionnelle d’O2 dans le peroxysome lors de l’oxydation du glycolate. Pour être complet, le NADH issu du fonctionnement de la glycine décarboxylase, membre du complexe enzymatique responsable de la transformation de glycine en sérine, peut être ré-oxydé par la chaîne respiratoire mitochondriale, conduisant à encore plus de consommation d’oxygène (voir aussi paragraphe 2.3).
2.3. La respiration mitochondriale
2.3.1. Approvisionnement de la mitochondrie en composés carbonés
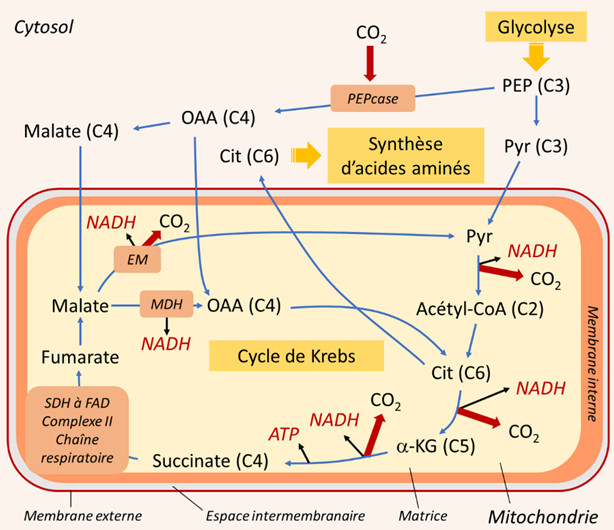
La respiration mitochondriale nécessite un approvisionnement de l’organite en composés carbonés issus des voies cytosoliques de dégradation des sucres. Deux voies se partagent cette tâche : la glycolyse et la voie des hexoses (sucres en C6) monophosphates (HMP). En fin de chaîne, la glycolyse donne le phosphoénolpyruvate (PEP) et le pyruvate (Pyr) (Figure 6) :
- Le pyruvate (en C3) est l’un des trois principaux acides organiques à pénétrer dans la mitochondrie.
- Le PEP (en C3) permet d’obtenir les deux autres (oxaloacétate et malate, acides organiques en C4) par une suite de deux réactions enzymatiques se déroulant dans le cytosol. La phosphoénolpyruvate carboxylase (PEPcase) permet d’obtenir, par carboxylation, de l’oxaloacétate (C4) qui donne ensuite du malate (C4).
La respiration mitochondriale proprement dite se décompose en deux parties :
- Le cycle de Krebs, qui se déroule dans la matrice des mitochondries ;
- La chaîne respiratoire, qui est intégrée dans la membrane interne des mitochondries.
2.3.2. Le cycle de Krebs
Le cycle de Krebs est alimenté par les trois acides organiques décrits précédemment (pyruvate, oxaloacétate et malate). Il est constitué d’une succession de réactions enzymatiques, dont certaines libèrent du CO2 et délivrent des produits énergétiques sous forme d’ATP et de NADH :
- L’une des enzymes participant au cycle de Krebs, la succinate déshydrogénase (SDH) est intégrée à la chaîne respiratoire dans le complexe II ;
- Lors de l’oxydation du succinate en fumarate, le co-enzyme FAD est réduit en FADH2 (Figure 7) ;
- Ce FADH2, ainsi que le NADH sont ensuite ré-oxydés par la chaîne respiratoire ;
- Le NADH est réduit soit au niveau du complexe I, soit par le biais d’une enzyme NADH déshydrogénase située sur la face interne de la membrane interne mitochondriale (NADin).
Quant au citrate, il peut sortir de la mitochondrie et servir de précurseur à la synthèse de composés aminés impliqués dans la résistance aux stress (Figure 6) (voir paragraphe 3.3).
2.3.3. La chaîne respiratoire ou chaîne de transfert d’électrons
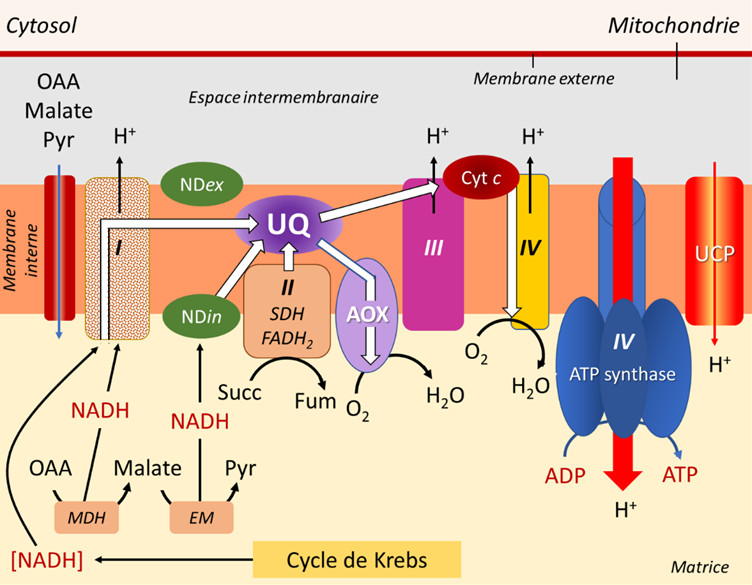
La chaîne respiratoire mitochondriale est constituée :
- D’une série de complexes respiratoires I, III et IV, reliés entre eux par des transporteurs d’électrons et de protons, prélevant les électrons fournis par le NADH et les amenant jusqu’à l’étape finale de réduction du dioxygène O2 en H2O (Figure 7) ;
- Le complexe II, quant à lui, fonctionne avec le FADH2;
- Les transporteurs impliqués sont essentiellement des cytochromes (petites protéines à Fer) et des ubiquinones (UQ, petites molécules à chaîne terpéniqueet mobiles au sein de la membrane lipidique) et la chaîne est souvent appelée voie cytochromique de transport d’électrons ;
- Au cours de ce cheminement, des protons sont rejetés dans l’espace intermembranaire à trois reprises, contribuant à l’établissement d’un gradient de protons ;
- Le retour des protons dans la matrice, canalisé par une ATPase (ou ATP synthase, lire Focus Synthèse d’ATP), permet la synthèse d’ATP (Figure 7).
Présence d’une oxydase alternative (AOX pour Alternative OXidase en anglais). Les plantes sont caractérisées par l’existence d’une respiration résistante au cyanure, particulièrement évidente lors de la thermogenèse chez les Aracées (Lire focus Thermogenèse et pollinisation chez les Aracées). Cette résistance au cyanure correspond au fonctionnement d’une double voie de transport mitochondrial d’électrons :
- La voie cytochromique, sensible au cyanure et couplée à la production d’ATP ;
- Une voie insensible au cyanure non-phosphorylante [3].
L’existence de ces deux voies permet de moduler la production d’ATP en fonction des besoins cellulaires en énergie [3].
Le pool d’ubiquinones fut identifié comme le point de branchement entre les deux voies de transfert d’électrons [4]. Le composé terminal de la voie de transfert d’électrons insensible au cyanure, dépendant d’une synthèse protéique nucléaire [5], est une quinol oxydase, appelée AOX (pour Alternative OXidase en anglais) [6].
La synthèse de l’oxydase alternative (AOX, pour Alternative OXidase en anglais) est (i) contrôlée par le noyau, de manière directe, et (ii) par le biais de signaux en provenance des mitochondries (signalisation rétrograde) et (iii) son expression est régulée de manière biochimique post-traductionnelle [7]. L’AOX peut prendre en charge une partie du transport d’électrons tout en réduisant la production globale d’ATP. Les contributions des complexes de la chaîne respiratoire et des deux oxydases terminales (cytochrome oxydase et alternative oxidase) au transport des électrons permettront d’ajuster la quantité d’ATP nécessaire aux besoins énergétiques.
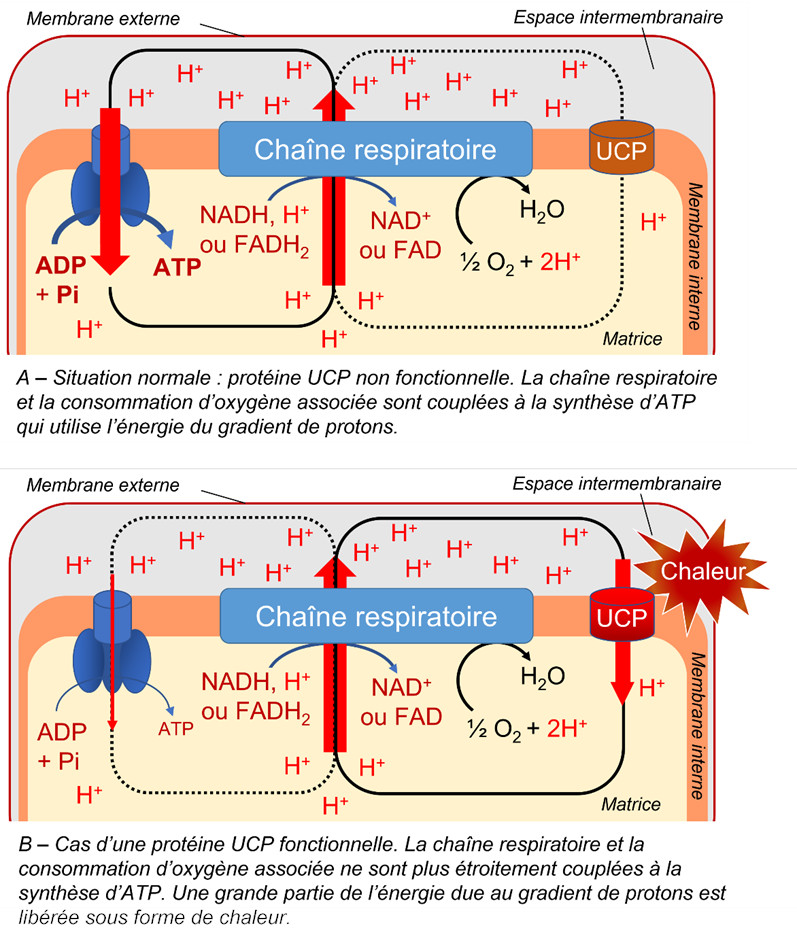
Présence d’une protéine de découplage. Découverte en 1995 dans les mitochondries végétales, la protéine de découplage (UCP pour UnCoupling Protein en anglais) :
- est insérée dans la membrane interne des mitochondries ;
- amoindrit la relation entre le gradient de protons lié au flux d’électrons et la synthèse d’ATP [9] (Figure 8) ;
- laisse repasser vers la matrice les protons libérés dans l’espace intermembranaire ;
- permet d’assurer un fort transport d’électrons sans synthèse concomitante d’ATP, ce qui peut se révéler intéressant lors de stress (voir paragraphe 3).
3. Rôle des processus respiratoires dans la réponse aux stress
3.1. La restitution de CO2 : perte ou nécessité ?
Les processus respiratoires (photorespiration et respiration) pourraient être perçus en termes de pertes de carbone. En effet, il est fait état fréquemment de pertes respiratoires. De fait, alors que la photosynthèse s’efforce d’assimiler un maximum de molécules de CO2 (opération bénéfique pour la diminution de l’effet de serre qui participe au dérèglement climatique), la respiration en relâche. Accessoirement, de l’oxygène est délivré lors de la photosynthèse (élément essentiel de la vie sur notre planète), alors qu’il sert d’oxydant dans les processus respiratoires. Cependant, deux raisons incitent à considérer la respiration de manière positive :
- Seulement un tiers des efforts assimilateurs de la photosynthèse sont dissipés lors de la respiration (voir paragraphe 2) ;
- Les processus respiratoires sont essentiels à la croissance des différents tissus de la plante en assurant trois objectifs :
- L’utilisation/transformation des produits initiaux de la photosynthèse (essentiellement des sucres) en squelettes carbonés qui permettront la synthèse de composés élaborés.
- La production d’énergie cellulaire (sous forme d’ATP), essentiellement au niveau des mitochondries, organites cellulaires respiratoires, nécessaire à la réalisation de ces synthèses.
- Le maintien d’un équilibre cellulaire redox en ajustant, toujours essentiellement au niveau mitochondrial, les rapports entre les formes réduites et oxydées des molécules de NADH et NADPH ainsi que la production d’ERO (espèces réactives de l’oxygène (Lire Contraintes environnementales et stress oxydant chez les plantes).
De plus, une augmentation de l’intensité respiratoire, parfois transitoire, est communément observée lors de stress subis par les plantes ou lors d’évènements réclamant une mobilisation importante de petits composés carbonés. Le phénomène de thermogénèse est emblématique de ce dernier cas [3] (Lire focus Thermogenèse et pollinisation chez les Aracées).

Lors d’un stress environnemental, l’équilibre entre production d’énergie et utilisation de composés carbonés pour la croissance est rompu. En effet, une partie des composés carbonés intermédiaires sont déviés vers la production de composés secondaires pour contrecarrer les dommages causés par le stress. Mais une augmentation de l’intensité respiratoire est-elle capable de satisfaire à l’ensemble des besoins (croissance et défense) ?
3.2. La photorespiration
La photorespiration, qui a lieu exclusivement le jour, joue un rôle significatif dans la réponse aux stress environnementaux.
- L’étape initiale du processus, qui met en compétition les deux fonctions (carboxylase et oxygénase) de la RubisCO, est très sensible au rapport entre les concentrations de CO2 et d’O2 dans l’environnement immédiat de la Rubisco. Deux contraintes au moins, toutes deux liées aux modifications actuelles des composantes climatiques, sont susceptibles d’augmenter le rapport O2/CO2 et, par la même, de causer une augmentation du cycle photorespiratoire.
- Une augmentation de température décroît à la fois la spécificité de la RubisCO pour le CO2 et la solubilité du gaz, conduisant à une plus forte pression d’O2 dans les cellules foliaires.
- La sécheresse, souvent liée à l’augmentation de la température, conduit à une fermeture des stomates, limitant ainsi l’entrée de CO2 et favorisant la fonction oxygénase, donc la photorespiration. La réduction de l’assimilation de CO2, liée à la diminution de la fonction carboxylase, réduit la consommation des composés réducteurs et de l’ATP issus de la chaîne de transport d’électrons photosynthétique. Cependant, celle-ci continue toujours de capturer l’énergie lumineuse avec un risque très important d’un excès d’électrons pouvant conduire à l’accumulation d’ERO et de NADPH.
- La photorespiration, en utilisant, au moins en partie, l’excès d’électrons lors de certaines de ses réactions enzymatiques, protège ainsi l’appareil photosynthétique de la photoinhibition. Elle est aussi capable de réalimenter le cycle de Calvin-Benson-Bassham en APG et en CO2, délivré dans la mitochondrie lors de l’oxydation de la glycine (voir Figure 5). La photorespiration pourrait ainsi contribuer à soulager une accumulation préjudiciable d’équivalents réduits, en parallèle avec des mécanismes de transfert spécifiques à l’évacuation de ce pouvoir réducteur excédentaire (présent dans NADPH) depuis le chloroplaste vers le cytosol et la mitochondrie.
3.3. Les voies de dégradation cytosoliques

- à l’apport de plus d’acides organiques (phosphoénolpyruvate, oxaloacétate, malate, pyruvate ; Figure 10) au contact de la mitochondrie ;
- au prélèvement de composés intermédiaires le long des voies dégradatives vers des voies de synthèse de composés secondaires liés à la défense contre les stress (flavonoïdes, phénylpropanoïdes…).
Un exemple intéressant est celui du pyruvate qui constitue le composé à 3 atomes de carbone emblématique de l’étape finale de la glycolyse (Figure 10). Le pyruvate, découvert par Berzélius en 1835 lors de la fermentation du raisin, dérive étymologiquement de l’association du grec pyro (feu) et du latin uva (raisin). Pyro, car cette molécule est brulée (on devrait dire oxydée) pour produire de l’énergie. Ce pyruvate provient essentiellement des sucres mais peut être produit par dégradation de lipides ou de protéines. Sa production à partir des sucres délivre de 20 à 30% de CO2 en plus par rapport aux deux autres sources [11].
Par ailleurs, le pyruvate est impliqué dans la stimulation de l’activité de l’AOX, ce qui représente un avantage en cas de stress (voir paragraphe 3.4.2).
Il faut ajouter que le citrate, premier acide organique formé dans le cycle de Krebs (voir Figure 6), peut être exporté de la mitochondrie vers le cytosol pour servir de précurseur à la synthèse de composés aminés impliqués dans la résistance aux stress [12].
3.4. Les mitochondries et la production d’ATP
Les acides organiques en provenance du cytosol vont être métabolisés dans la matrice mitochondriale au niveau du cycle de Krebs, appelé aussi cycle des acides tricarboxyliques (voir Figure 6). Du CO2 et du pouvoir réducteur (essentiellement sous forme de NADH) sont produits à cette occasion. La réoxydation des composés réduits se fait en présence d’oxygène grâce à la chaîne respiratoire insérée dans la membrane interne de la mitochondrie et permet la production d’énergie sous forme d’ATP (voir Figure 7). Mais le fonctionnement de cette chaîne respiratoire s’accompagne inévitablement de la production d’Espèces Réactives de l’Oxygène (ERO), production qu’il faut réguler d’autant plus que les stress entraînent une production accrue de ces ERO [12] (Lire Contraintes environnementales et stress oxydant chez les plantes).
Les mitochondries vont donc présenter toute une gamme de solutions pour répondre au mieux aux contraintes subies par les plantes, au moins lors de stress d’intensité moyenne ou s’exerçant sur des durées modérées.
3.4.1. Les différentes voies de transport d’électrons
Les électrons fournis par les composés réduits de type NADH (parfois aussi NADPH), produits dans la matrice mitochondriale par le cycle de Krebs et les enzymes associées, sont amenés à parcourir une chaîne d’oxydoréduction impliquant un grand nombre de transporteurs d’électrons et parfois de protons, dont certains imbriqués dans des complexes (I, III et IV) (voir Figure 7).
- Les complexes I, III et IV sont effectivement associés à l’émission de protons dans l’espace intermembranaire.
- Quant aux électrons transmis par le FADH2, ils sont spécifiquement pris en charge par le complexe II qui, lui, n’est pas associé à un transfert de protons (Figure 7).
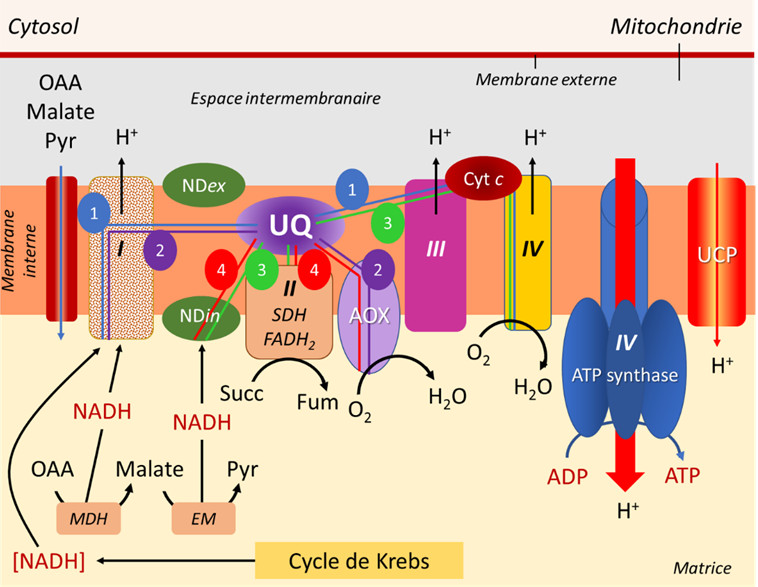
- La voie 1 (en bleu), la plus énergisante, passe par tous les complexes et permet l’obtention de 3 ATP.
- La voie 2 (en violet) utilise le complexe I mais court-circuite les 2 autres complexes en utilisant l’AOX, ce qui conduit à la production d’1 ATP
- La voie 3 (en vert) court-circuite le complexe I grâce au NDin, puis utilise les complexes III et IV, ce qui permet d’obtenir 2 ATP.
- La voie 4 (en rouge) court-circuite tous les complexes en passant par NDin et AOX, ne permettant aucune production d’ATP.
- Quant aux électrons en provenance du FADH2, leur parcours peut inclure les deux derniers complexes III et IV, permettant l’obtention de 2 ATP, ou utiliser seulement l’AOX, conduisant à 0 ATP.
Cette plasticité est déjà un atout intéressant permettant de moduler la quantité d’énergie nécessaire pour le fonctionnement harmonieux des cellules végétales dans des conditions environnementales favorables :
- En cas de stress, la répartition des électrons dans les différentes voies pourra évoluer afin de privilégier la combinaison adaptée à la nouvelle demande énergétique.
- La production d’ERO au niveau des complexes I, II et III, usuelle dans des conditions favorables, augmente systématiquement lors d’un stress, ce qui rend intéressant l’utilisation de voies privilégiant des transporteurs court-circuitant ces complexes (voir 3.4.2 et 3.4.3).
3.4.2. L’alternative oxydase (AOX)
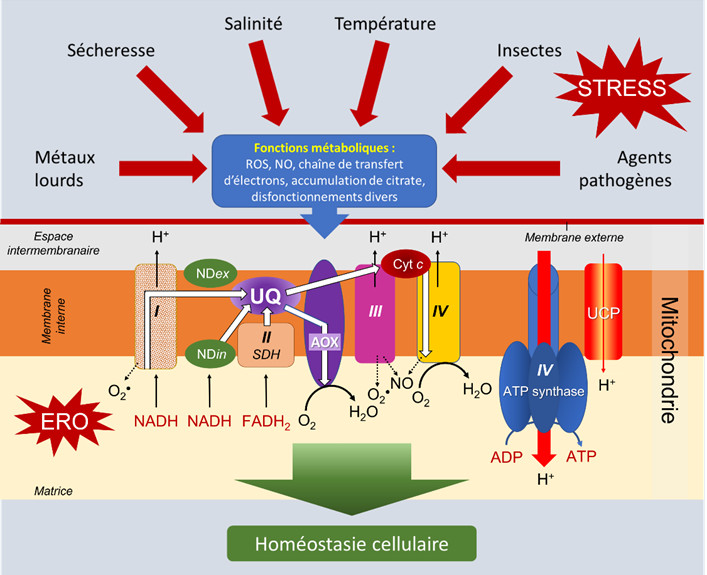
L’oxydase alternative, dite non phosphorylante, surtout associée à la NADH déshydrogénase interne (NDin) qui court-circuite le complexe I, permet de réduire considérablement la production d’ERO. L’augmentation de son implication dans le transport d’électrons lors de stress a été démontrée [8], [12] (Lire Contraintes environnementales et stress oxydant chez les plantes).
Le fonctionnement de l’AOX est en effet stimulé par la présence de plus abondantes quantités de pyruvate lors de stress et des effecteurs signalent au noyau la nécessité d’initier la synthèse de l’enzyme mitochondriale. La fonction dévolue à l’AOX en réponse à des contraintes environnementales semble donc particulièrement importante (Figure 12) [13].
3.4.3. La protéine découplante (UCP)
Toujours parce qu’elle évite une sur-production d’ATP, la protéine découplante voit son importance augmenter lors de stress [14]. Les protons de l’espace intermembranaire repassent dans la matrice en court-circuitant l’ATP synthétase. La diminution du potentiel de membrane diminue alors fortement la production d’ERO sans affecter la production indispensable d’énergie. La protéine découplante participe ainsi, à côté de l’AOX, à l’amenuisement de la présence potentiellement toxique de ces ERO dans les mitochondries et les cellules [15].
4. Messages à retenir
- La respiration et la photosynthèse sont des processus complémentaires participant à la croissance équilibrée des plantes. L’utilisation abusive du terme « perte respiratoire » doit donc être évitée.
- Les processus respiratoires comprennent la photorespiration et la respiration sensu stricto, parfois dénommée « mitochondriale ». La photorespiration se déroule le jour et fait intervenir cycliquement chloroplastes, peroxysomes et mitochondries. La respiration, quant-à-elle, a lieu jour et nuit, et fait intervenir les processus dégradatifs localisés dans le cytosol (glycolyse et voie des pentoses phosphates) et la mitochondrie.
- Environ un tiers de l’assimilation photosynthétique de CO2 est utilisé par les processus respiratoires. Ceux-ci jouent essentiellement un double rôle de production d’ATP (respiration totale jusqu’à l’émission de CO2) et d’utilisation de composés carbonés intermédiaires pour réaliser les synthèses des composés structuraux nécessaires à la croissance.
- Des stress environnementaux se traduisent par une augmentation des processus respiratoires avec synthèse accrue de composés carbonés nécessaires à la défense de la plante. Ajoutée à une diminution de l’assimilation photosynthétique, cette nouvelle répartition des ressources disponibles pourrait conduire, à moyen terme, à un déséquilibre fâcheux pour la croissance. De plus, les stress occasionnent une augmentation de la formation d’espèces réactives de l’oxygène (ERO).
- Afin de réduire les risques de toxicité engendrés par une production d’ERO trop élevée, des mécanismes sont mis en œuvre pour privilégier des systèmes mitochondriaux qui affaiblissent la production d’ATP et réduisent l’utilisation des complexes producteurs d’ERO (AOX, UCP, NDin).
- Ceci ne fonctionne que si le stress n’est pas trop intense ou est de courte durée. Dans le cas contraire, la diminution de plus en plus marquée des ressources carbonées issues de l’assimilation, conjuguée à une demande croissante d’approvisionnement de composés de défense, conduirait inexorablement à une réduction de plus en plus forte de la croissance.
Notes & References
Image de couverture. Paysage pris depuis le chemin des cimes vers Drachenbronn-Birlenbach, Parc naturel régional des Vosges du Nord [Source : Photo © P. Dizengremel, août 2021]
[1] Bediee A, Luchaire N, Dambreville A, Brichet N, Hamard P, Dauzat M, Muller B & Pellegrino A (2015). Mesures en série des échanges gazeux à l’échelle plante entière de plantes cultivées en pot. Cahier des Techniques de l’INRA, 86, 21 p. hal-01329223
[2] Möller CM, Müller D, & Nielsen J (1954). Graphic presentation of dry matter production of European beech. Det Forstlige Forsogsvoesen i Danmark, XXI, 327-335.
[3] Meeuse BJD (1975). Thermogenic respiration in aroids. Annu. Rev. Plant Physiol., 26, 117-126.
[4] Storey BT (1976). Respiratory Chain of Plant Mitochondria. XVIII. Point of interaction of the alternative oxidase with the respiratory chain. Plant Physiol., 58, 521-525.
[5] Dizengremel P & Lance C (1976). Control of changes in mitochondrial activities during aging of potato slices. Plant Physiol., 58, 147-151.
[6] Hiser C & McIntosh L (1990). Alternative oxidase of potato is an integral membrane protein synthesized de novo during aging of tuber slices. Plant Physiol., 93, 312-318.
[7] Vanlerberghe GC (2013). Alternative oxidase: a mitochondrial respiratory pathway to maintain metabolic and signaling homeostasis during abiotic and biotic stress in plants. Int. J. Mol. Sci., 14, 6805-6847; doi:10.3390/ijms14046805.
[8] Selinski J, Scheibe R, Day D & Whelan J (2018). Alternative oxidase is positive for plant performance. Trends in Plant Science, 23, 588-597; https://doi.org/10.1016/j.tplants.2018.03.012.
9 Vercesi AE, Borecký J, de Godoy Maia I, Arruda P, Cuccovia IM, & Chaimovich H (2006). Plant uncoupling mitochondrial proteins. Annu. Rev. Plant Biol., 57, 383-404. doi: 10.1146/annurev.arplant.57.032905.105335.
[10] Libérez Psikoda ! Journal La Hulotte N° 26, 1 janvier 1985 ; https://www.lahulotte.fr/
[11] Le XH, Lee CP, Monachello D, & Millar AH (2022). Metabolic evidence for distinct pyruvate pools inside plant mitochondria. Nature Plants, 8, 694-705; https://doi.org/10.1038/s41477-022-01165-3.
[12] Gandin A, Dizengremel P, & Jolivet Y (2021). Integrative role of plant mitochondria facing oxidative stress : the case of ozone. Plant Physiol. Biochem., 159, 202-210. https://doi.org/10.1016/j.plaphy.2020.12.019.
[13] Saha B, Borovskii G, Kumar Panda S (2016). Alternative oxidase and plant stress tolerance. Plant Signaling and Behavior, 11, e1256530-1 – e1256530-4. http://dx.doi.org/10.1080/15592324.2016.1256530
[14] Barreto P, Counago RM, & Arruda P (2020). Mitochondrial uncoupling protein-dependent signaling in plant bioenergetics and stress response. Mitochondrion, 53, 109-120. https://doi.org/10.1016/j.mito.2020.05.001.
[15] Borecky J, & Vercesi AE (2005). Plant uncoupling mitochondrial protein and alternative oxidase: energy metabolism and stress. Bioscience Reports, 25, 271-286. DOI: 10.1007/s10540-005-2889-2.
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Pour citer cet article : DIZENGREMEL Pierre (10 mai 2023), Résistance des plantes aux stress : rôle de la respiration ?, Encyclopédie de l’Environnement. Consulté le 23 avril 2025 [en ligne ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/vivant/resistance-plantes-stress-role-respiration/.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.


