



Symbiose et parasitisme
PDF
Aucun organisme ne vit seul. Il est en permanence associé plus ou moins étroitement à de nombreux autres organismes, microorganismes en particulier. Leurs interactions peuvent être classées en fonction du degré d’association des organismes impliqués, de la durée de ces interactions et de leur caractère bénéfique (ou non) pour l’un et l’autre des partenaires. Toutes les situations intermédiaires existent, formant un véritable continuum des organismes libres qui ont besoin d’autres organismes pour se nourrir jusqu’aux parasites dont le cycle de vie repose entièrement sur des hôtes bien spécifiques. Symbiose et parasitisme permettent d’illustrer que, au-delà de l’extrême diversité des situations, l’interaction est dans tous les cas essentielle à la vie des partenaires, et souvent à l’origine de l’émergence de propriétés nouvelles des systèmes ainsi constitués. C’est par exemple le cas des microbiotes associés à chacun des êtres vivants. Mais c’est aussi le cas d’organismes modifiés par les parasites qui les infectent et vont jusqu’à perturber le comportement des hôtes contaminés en comparaison des individus sains.
1. Quels types d’interactions entre organismes vivants ?
Le réseau d’interactions et d’interdépendances qui existe entre des milliards d’organismes au sein de la biosphère est un niveau d’organisation du Vivant fondateur du concept de biodiversité (lire Qu’est-ce que la biodiversité ?). Ces interactions sont le plus souvent à bénéfices réciproques et leur rôle dans la physiologie et l’adaptation des organismes est essentiel. Par exemple, bien des animaux ne digèrent pas sans l’aide des bactéries de leur tube digestif, la plupart des plantes ne peuvent exploiter le sol qu’à l’aide de champignons colonisant leurs racines, qu’elles nourrissent en retour [1].
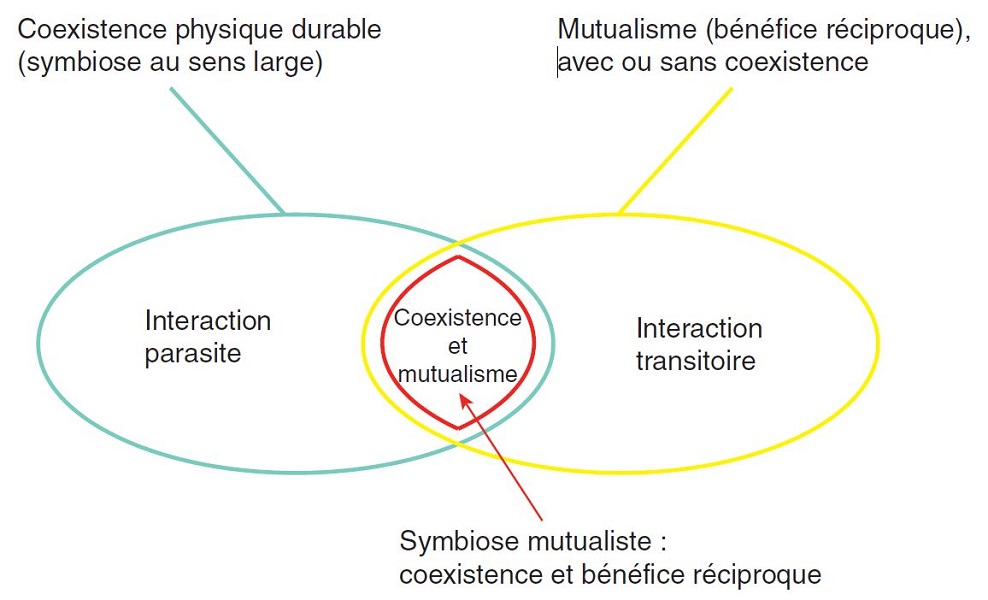
Mais il n’en est pas toujours ainsi : les interactions entre deux organismes peuvent être classées en fonction de leur caractère bénéfique, néfaste ou neutre pour l’un et l’autre des partenaires. Ainsi, peuvent être distinguées :
– des interactions bénéfiques pour l’un des partenaires et néfastes pour l’autre (prédation, parasitisme),
– des interactions bénéfiques pour l’un et neutres pour l’autre (commensalisme),
– et des interactions à bénéfices réciproques (mutualisme).
De plus, toutes les situations intermédiaires existent, en un véritable continuum de types d’interaction (Figure 1) [2]. Elles peuvent également être classées en fonction de leur caractère instantané (prédation) ou durable (parasitisme, mutualisme, etc.), ainsi que d’après le degré d’association entre les partenaires [2].
Étymologiquement, le terme de symbiose désigne « la vie en commun d’organismes d’espèces distinctes ». Cette définition large désigne une coexistence durable, impliquant tout ou partie du cycle de vie des deux organismes, quels que soient les échanges entre ceux-ci. Une définition plus restrictive réserve le terme de symbiose aux coexistences durables et mutualistes (partie rouge sur la Figure 1).
2. Les symbioses mutualistes


La symbiose peut protéger contre les agressions du milieu, en particulier quand un des partenaires vit à l’intérieur de l’autre. Dans les mycorhizes, le champignon est souvent protégé dans la racine (où il stocke ses réserves, dans le cas de certaines endomycorhizes), mais il peut aussi protéger la racine lorsqu’il forme un manchon autour de la racine (ectomycorhizes). Un champignon du genre Neotyphodium (Figure 2E) vit en symbiose à l’intérieur de la fétuque élevée (Festuca arundinacea) qu’il protège des herbivores en secrétant des alcaloïdes toxiques pour les insectes et les mammifères. Ces derniers évitent alors les zones herbeuses ainsi protégées. De son coté, le champignon se propage de génération en génération en colonisant les graines [4].
La protection est parfois le seul bénéfice obtenu, comme dans les symbioses nettoyeuses des écosystèmes coralliens, où un petit animal (poisson ou crevette) nettoie la peau et/ou les cavités de l’autre, éliminant des débris et des petits parasites (Figure 3). Cependant, certaines anémones de ces récifs grandissent plus vite, ont de plus grandes chance de survie et présentent une plus grande densité de Zooxanthelles (algues symbiotiques unicellulaires vivant au sein des anémones de mer) lorsqu’elle sont fréquentées par des poissons. Cette augmentation des performances des anémones est le fruit d’un transfert de nutriments du poisson à l’anémone, qui exploite l’urine du premier comme source d’azote et de phosphate [5]. Ces observations démontrent que les poissons-clowns contribuent à nourrir l’anémone de mer et que les algues symbiotiques de celles-ci bénéficient de cet apport [6].
D’autres bénéfices reposent sur la capacité d’un des partenaires à se déplacer (pollinisation par les abeilles, dispersion des graines par les fourmis ou les oiseaux). Au bilan, des associations au fonctionnement semblable ont été mises en place plusieurs fois au cours de l’évolution. De telles convergences sont illustrées par la diversité des insectes cultivant des champignons (fourmis, termites, coléoptères) et des eucaryotes abritant des algues photosynthétiques dans leurs cellules (comme l’apparition des chloroplastes au sein de la cellule eucaryote) (lire Symbiose et évolution). Tous les organismes ont eu l’occasion de contracter, au cours de leur évolution, une ou plusieurs symbiose(s) mutualiste(s). C’est en particulier vrai pour des organismes multicellulaires de grande taille, qui constituent un écosystème pour les organismes microscopiques. La rhizosphère (le sol entourant la racine des plantes) ou le tube digestif des animaux sont ainsi des niches microbiennes majeures, peuplées de milliers d’espèces pour chaque individu-hôte, dont certains occupants sont favorables à l’hôte. En conséquence, chaque organisme possède un cortège de symbiotes, surtout développé chez les multicellulaires.
3. Propriétés émergentes de la symbiose
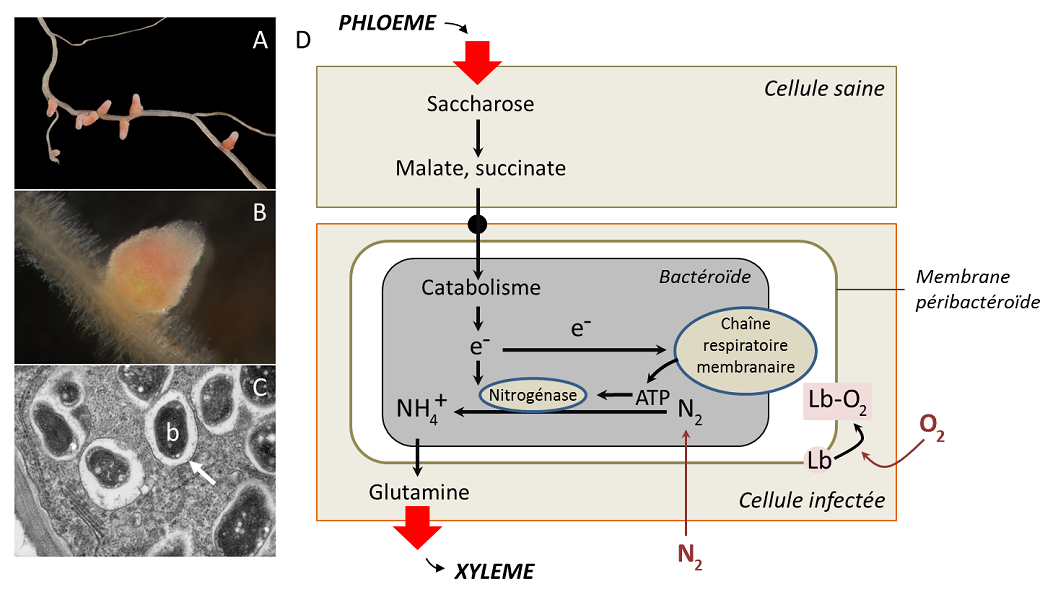
D’autres émergences sont fonctionnelles. Dans l’exemple des nodosités (Figure 4 D), le bactéroïde utilise de l’énergie qu’il obtient de sa respiration pour réduire, grâce à la nitrogénase, l’azote atmosphérique en ammonium, qui sert de source d’azote à la plante (et au bactéroïde). Réciproquement, la plante lui fournit du carbone et un approvisionnement en oxygène. Il faut de l’oxygène pour la respiration, mais la nitrogénase est inactivée par l’oxygène : cette contradiction explique qu’un rhizobium libre dans le sol est incapable de fixer de l’azote. En revanche, dans la nodosité, l’oxygène ne diffuse pas librement, mais est capté par une protéine de la cellule-hôte, la leghémoglobine [7]. Située autour du bactéroïde, la leghémoglobine protège la nitrogénase des effets de l’oxygène qui l’inactive et constitue une réserve d’oxygène pour la respiration des bactéries. La fixation de l’azote n’est donc réalisée que dans la nodosité (lire Des plantes qui vivent de l’air du temps).
De nombreux autres traits fonctionnels sont induits par la symbiose, comme certains effets protecteurs qui reposent sur l’induction de défenses du partenaire, tolérées par le symbiote mais nocives pour des pathogènes. Les champignons mycorhiziens induisent par exemple l’accumulation de tanins protecteurs au niveau racinaire qui sont responsables de l’induction d’un niveau de défense et de réactivité accru dans toute la plante, parties aériennes comprises. Ainsi, la plante mycorhizée réagit plus vite et plus fortement à un herbivore ou un parasite qu’une plante contrôle non-mycorhizée. Chez les lichens, l’algue induit la synthèse par le champignon de métabolites secondaires qui ont un rôle protecteur contre les forts éclairements et contre les herbivores.
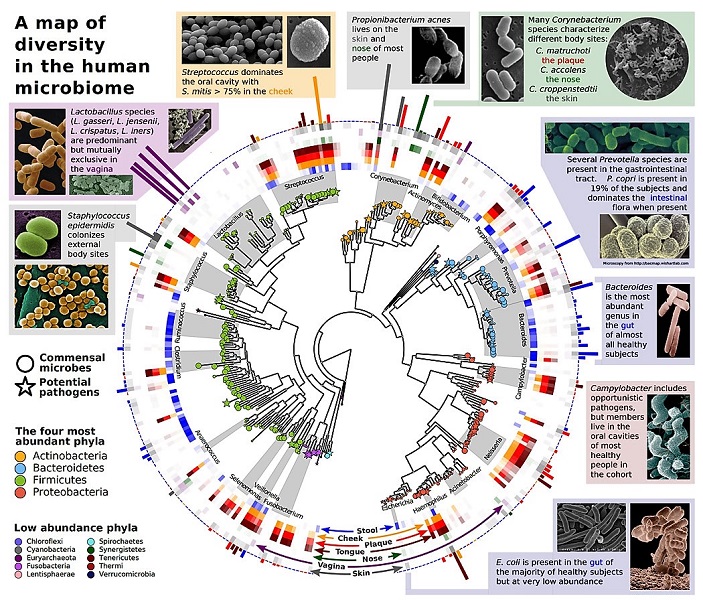
Le microbiote intestinal est indispensable au bon fonctionnement de son hôte humain, non seulement au niveau de la digestion ou de la fabrication de vitamines, bien sûr, mais aussi pour le métabolisme, l’immunité… ou bien encore pour le système nerveux. Les déséquilibres de la flore intestinale sont aujourd’hui suspectés d’être à l’origine d’une kyrielle de pathologies : obésité, diabète, maladies cardiovasculaires, allergies, maladies inflammatoires, voire l’autisme [2],[7]. Les microbiotes humains ne se limitent pas au tube digestif : les programmes internationaux de métagénomique ont permis d’identifier les gènes provenant de très nombreux microrganismes symbiotiques vivant dans la bouche, le nez, le vagin ou sur la peau (Figure 5) (lire Les microbiotes humains : des alliés pour notre santé).
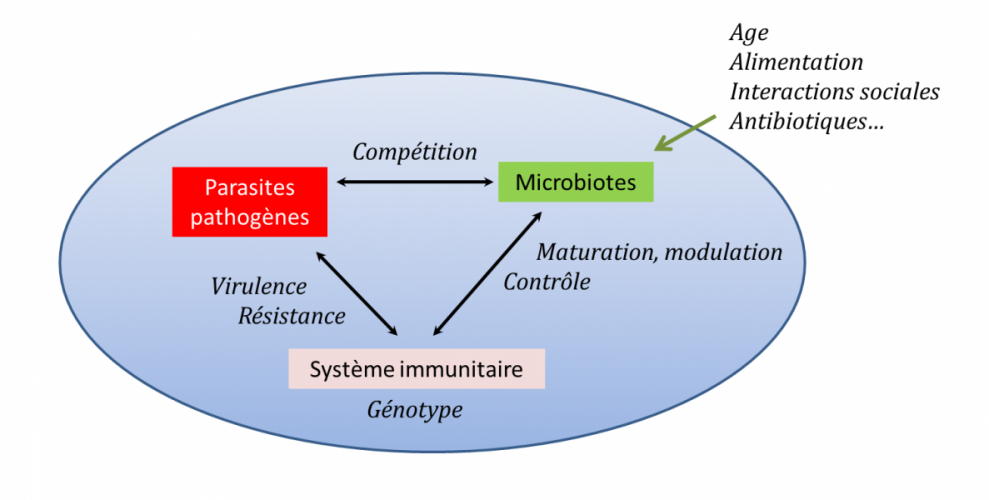
Il a donc été proposé que l’unité pertinente pour la biologie ou pour l’évolution soit moins l’organisme que le cortège symbiotique : on parle d’holobionte pour dénommer cette entité plus pertinente au regard de l’importance des interactions biotiques [11].
4. Le parasitisme, une réussite évolutive
Si l’un des partenaires de la symbiose découvre le moyen d’utiliser efficacement l’autre, il en devient alors un parasite. Il existe en effet un continuum entre symbiose et parasitisme [5]. Le parasite exploite des ressources fournies par un autre individu non apparenté, l’hôte, au détriment de celui-ci. Le parasitisme est une interaction durable avec un hôte, contrairement à la prédation au cours de laquelle l’interaction ne dure guère que le temps de la capture et de la digestion. Cela dit, d’un point de vue évolutif, on peut dire que la prédation n’est qu’une forme extrême de parasitisme. Il existe des parasites tuant lentement leur hôte. C’est le cas de champignons parasites de plantes (Mildiou, Armillaires, Amadouviers, etc…) qui achèvent leur cycle de vie sur les tissus morts. Lorsqu’un guépard se saisit d’une antilope, il y a échange d’énergie et d’énergie seulement. Dans les systèmes parasites-hôtes où l’hôte survit (on parle de parasitisme biotrophe), la durée de l’interaction est tout autre : les deux organismes vivent alors ensemble, souvent l’un dans l’autre, parfois cellule dans cellule ou même génome dans génome. Les informations génétiques de chacun des partenaires s’expriment ainsi côte à côte et durablement dans une portion d’espace minuscule [11].
Tous les êtres vivants sont concernés par le parasitisme en tant qu’hôte ou parasite (Figure 7). Parmi les espèces connues, 30 % des quelques 2 millions d’espèces eucaryotes seraient des parasites [12]. La parasito-faune la mieux connue est celle de l’homme. Elle consiste en 179 espèces de parasites, parmi lesquelles 35 paraissent spécifiques d’Homo sapiens [13]. Cette image peut être aggravée par l’hyperparasitisme (parasites de parasites), un phénomène répandu chez les arthropodes parasites et parasitoïdes [14]. Des estimations récentes suggèrent que le monde des virus, qui parasitent les cellules en détournant leur fonctionnement vers la production de nouvelles particules virales, a été profondément sous-estimé. Ils sont présents dans tous les écosystèmes et constitueraient les entités génétiques les plus abondantes et diversifiées du vivant [15].
- Vidéo « Comment le SARS-CoV-2 transforme une cellule en usine à virus » (© 2020 Diffusonslascience.fr)
Pour les évolutionnistes, les modèles hôtes-parasites suscitent d’innombrables questions, relatives au parasitisme lui-même, aux dynamiques évolutives de leurs interactions, ainsi qu’aux conséquences évolutives sur les espèces hôtes. Le rôle joué par les parasites sur le monde des espèces libres est en effet énorme. Le succès du mode de vie parasitaire ne s’est jamais démenti pendant toute la durée de l’évolution car un hôte offre, à tout organisme qui sait l’exploiter, non seulement habitat et nourriture mais aussi un moyen efficace de dispersion. Si, dans le passé, les travaux ont surtout considéré les effets directs des pathogènes sur la fécondité et la survie de leurs hôtes, les recherches actuelles illustrent des conséquences sur des traits aussi divers que le comportement, les processus de sélection ou l’histoire de vie, pour ne citer qu’eux.

5. Les cycles parasitaires
Le cycle parasitaire est la suite des transformations subies par un parasite au cours de sa vie pour assurer sa reproduction, dans les diverses niches écologiques qu’il occupe : hôte(s), milieu extérieur. Si de nombreuses espèces de parasites ont des cycles simples, exploitant une seule espèce d’hôte, d’autres exploitent successivement plusieurs espèces d’hôtes : cela permet des relais saisonniers, ou de multiplier les formes infectieuses, car le taux de succès de colonisation des hôtes est souvent faible. La complexification des cycles est apparue plusieurs fois indépendamment au cours de l’évolution. Parmi les records de complexité, on peut citer le cas du trématode Halipegus ovocaudatus dont le cycle comporte 4 hôtes obligatoires : un mollusque, un crustacé copépode, une larve de libellule et une grenouille. A côté de ces situations extrêmes, on trouve des cycles complexes à deux ou trois hôtes, notamment chez les helminthes ou les rouilles (des champignons pathogènes). À côté des complexifications de cycles simples, il y a également au cours de l’évolution des simplifications secondaires de cycles complexes.
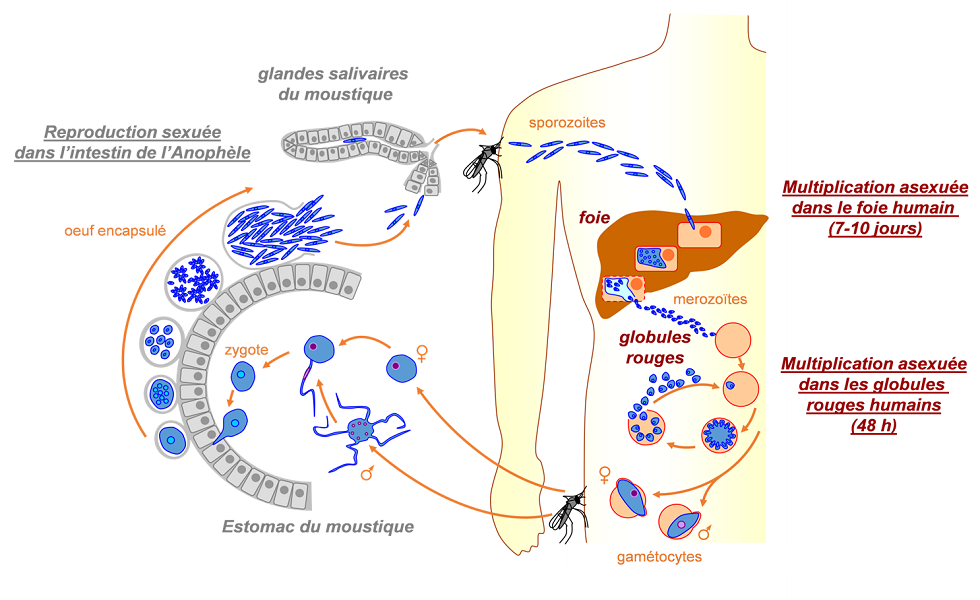
Lorsqu’ils sont infectés par plasmodium, les moustiques changent de comportement : ils deviennent plus actifs, plus agressifs et piquent plus de personnes, augmentant ainsi leur probabilité de transmission [2]. Ces changements semblent être synchronisés avec le développement du parasite (e.g. diminution du taux de piqûres des moustiques lorsque le parasite est immature, et augmentation lorsque le parasite a rejoint le stade transmissible). Une fois chez l’hôte vertébré, ces mêmes parasites semblent capables de modifier les odeurs des hôtes afin de les rendre plus attractifs pour les moustiques vecteurs. Cette modification du comportement des hôtes après infection est un exemple caractéristique de manipulation parasitaire [2].
6. Les manipulations parasitaires
Certains parasites sont en effet capables de modifier de façon notable la physiologie, la morphologie ou encore le comportement de leur hôte avec pour conséquence l’augmentation de leur probabilité de transmission. Cette stratégie d’exploitation de l’hôte est à présent décrite dans de nombreux systèmes hôtes-parasites phylogénétiquement éloignés. Les modifications phénotypiques chez les hôtes infectés sont généralement considérées comme une illustration du concept de phénotype étendu [18]. Ces changements phénotypiques correspondent en fait à l’expression des gènes du parasite et à l’effet des protéines correspondantes sur le phénotype de l’hôte. Selon cette idée, ces modifications induites sont adaptatives pour le parasite et non pour l’hôte.

- Après que la larve de la guêpe parasitoïde Dinocampus coccinellae soit sortie de l’abdomen de la coccinelle parasitée, puis se soit transformée en cocon, la coccinelle va s’en occuper et protéger le cocon jusqu’à l’émergence de la guêpe (Figure 9 A).
- La guêpe Glyptapanteles pond des d’œufs à l’intérieur de la chenille du papillon Thyrinteina leucocerae qui va changer de comportement après l’éclosion des œufs et leur transformation en pupe : elle arrête de se nourrir, devient immobile, et protège les pupes contre les prédateurs jusqu’à leur éclosion (Figure 9 C).
Certaines manipulations sont encore plus extrêmes :
- En envahissant une fourmi de la forêt brésilienne, le champignon Ophiocordyceps camponoti-rufipedis (Figure 9 B) en manipule le comportement en prenant le contrôle de son « cerveau », conduisant la fourmi à grimper au sommet d’une plante où les conditions (lumière, humidité) sont favorables au développement du champignon. Une fois la fourmi fermement cramponnée à la tige, le champignon la tue et prolifère tranquillement : les spores produites sont alors facilement dispersées au gré du vent.
- Une stratégie un peu semblable est mise en œuvre dans le cas de l’escargot Succinea putris infecté par le parasite Leucochloridium paradoxum. Ce dernier se loge dans les antennes de l’escargot qui vont prendre l’aspect et les mouvements d’un ver, devenant une proie d’autant plus repérable pour les oiseaux que le comportement de l’escargot est aussi modifié car il a tendance à quitter la protection de la végétation. Le cycle de vie du parasite se poursuit dans l’oiseau qui a mangé l’escargot et dont les fientes permettent la dissémination des œufs du parasite (Figure 9 E).
- De son côté, la sacculine (Sacculina carcini), un petit crustacé parasite du crabe, colonise son hôte, modifie son équilibre hormonal et l’empêche de se reproduire, sa seule fonction étant de nourrir le parasite (Figure 9 F). Après la fécondation de la sacculine, le crabe s’occupera des œufs du parasite comme si c’était les siens…
Certaines manipulations par des parasites conduisent l’hôte à avoir un comportement suicidaire. Un cas bien décrit est celui de vers nématomorphes, dont la forme adulte vit dans l’eau et ressemble à une sorte de fil. L’hôte est en général un insecte terrestre, par exemple une sauterelle (Figure 7 H) qui héberge la forme larvaire. Le ver devenu adulte doit rejoindre le milieu aquatique pour se reproduire. Pour cela, il manipule le comportement de l’hôte, obligeant ce dernier à se jeter dans l’eau. Il peut alors, grâce cette noyade finale, rejoindre le milieu dans lequel il boucle son cycle de vie. Ce type de comportement suicidaire peut être néanmoins avantageux pour les semblables d’un hôte infecté, car il permet de réduire les risques de contamination. C’est le cas de la fourmi parasitée par Ophiocordyceps (figure 9B) qui est alors reconnue comme telle et rejetée de la fourmilière par ses congénères.
Dans quelques cas concernant les végétaux, le déterminisme de la manipulation est un peu mieux connu. Il révèle un mécanisme étonnamment convergent chez des champignons et des bactéries pathogènes, mais aussi des nématodes parasites de plantes. Ils provoquent des déformations racinaires où ils s’abritent se nourrissent, ce qu’on appelle des galles. Les génomes de ces organismes codent une multitude de petites protéines (ou peptides) sécrétées, qui modifient le fonctionnement d’autres protéines de l’hôte. On parle d’effecteurs : certains pénètrent dans les cellules de l’hôte, et réorganisent le métabolisme ou altèrent les réactions de défense… Parfois, ils agissent au niveau du noyau de la cellule et sont responsables de modifications de l’expression génétique. Il est probable que ces mécanismes jouent aussi dans d’autres types de parasitisme : on les retrouve même chez des champignons mycorhiziens. Ceci suggère que les peptides sécrétés pourraient contribuer aux modifications observées dans les symbioses mutualistes – soulignant une fois encore l’existence de similitudes de mécanismes entre symbiose mutualiste et parasitisme.
Au-delà de l’aspect spectaculaire et fascinant que recouvre les manipulations parasitaires, certains des pathogènes impliqués sont à l’origine de nombreuses pertes de récoltes, mais aussi de graves maladies, notamment des maladies à transmission vectorielle telles que le paludisme évoqué plus haut, la dengue, les trypanosomiases ou les leishmanioses, et représentent ainsi des problèmes majeurs de santé publique [19].
Références et notes
Photo de couverture : © Jenny Huang from Taipeh (CC BY 2.0) via Wikimedia Commons
[1] Selosse M.A. (2000). La Symbiose. Vuibert, Paris.
[2] Lefèvre T., Renaud F., Selosse M.-A., Thomas F. (2010). Chapitre 14, Évolution des interactions entre espèces, in F. Thomas, T. Lefèvre & M. Raymond (ed.), Biologie évolutive, p. 555-653. De Boeck, Paris.
[3] Selosse MA & Roy M (2012) Les plantes qui mangent des champignons… Dossier Pour la Science « Les végétaux insolites » 77: 102-107
[4] Selosse MA, Gilbert A (2011) Des champignons qui dopent les plantes. La Recherche 457, 72-75.
[5] Cleveland A., Verde E.A. & Lee R.W. (2011) Nutritional exchange in a tropical tripartite symbiosis: direct evidence for the transfer of nutrients from anemonefish to host anemone and zooxanthellae, Marine Biology, 158: 589–602
[6] Corbara B http://www.especes.org/#/1-menage-a-trois/4541755
[7] Leghémoglobine. Protéine fixatrice de dioxygène, de structure très proche de l’hémoglobine du sang. Présente dans les nodosités des légumineuses,elle protège le complexe enzymatique (nitrogénase/hydrogénase) des effets du dioxygène qui l’inactive et constitue une réserve d’oxygène pour les bactéries (activité aérobie).
[8] Dawkins R. (1982) The extended phenotype. Oxford University Press, Oxford.
[9] Morgan X.C., Segata N. & Huttenhower C. (2013) Biodiversity and functional genomics in the human microbiome, Trends Genet. 29, 51–58
[10] Gross R., Vavre F., Heddi A., Hurst G.D.D., Zchori-Fein E. &Bourtzis K. (2009) Immunity and symbiosis. Molecular Microbiology 3, 751-759.
[11] Selosse M.A. (2016) Au delà de l’organisme : l’holobionte. Pour la Science, 269, 80-84.
[11] Combes C. (1995) Interactions durables. Écologie et évolution du parasitisme. Éditions Masson, 525 p.
[12] De Meeûs T. & Renaud F. (2002). Parasites within the new phylogeny of eukaryotes.Trends in Parasitology 18, 247-251.
[13] De Meeûs T., Prugnolle F. & Agnew P. (2009). Asexual reproduction in infectious diseases. In Lost Sex, Schön I, Martens K & van Dijk P eds, Springer, NY., p. 517-533.
[14] Parasitoïde : organisme qui se développe sur ou dans un organisme « hôte » dans un processus à deux phases : il est tout d’abord biotrophe puis prédateur, ce qui conduit à la: mort finale de l’hôte.
[15] Hamilton G. (2008). Welcome to the virosphere.New Scientist 199, 38-41.
[16] Source http://www.archive.org/stream/traitzoologiqu00brem#page/n613/mode/2up
[17] Zheng et al. (2011) Genome sequence of the insect pathogenic fungus Cordyceps militaris, a valued traditional Chinese medicine. Genome Biology 12: R116
[18] Dawkins R. (1976) The selfish gene. Oxford University Press.
[19] Lefèvre T. & Thomas F. (2008) Behind the scene, something else is pulling the strings: Emphasizing parasitic manipulation in vector-borne diseases. Infection, Genetics and Evolution 8, 504–519.
L’Encyclopédie de l’environnement est publiée par l’Association des Encyclopédies de l’Environnement et de l’Énergie (www.a3e.fr), contractuellement liée à l’université Grenoble Alpes et à Grenoble INP, et parrainée par l’Académie des sciences.
Pour citer cet article : SELOSSE Marc-André, JOYARD Jacques (11 février 2022), Symbiose et parasitisme, Encyclopédie de l’Environnement. Consulté le 5 avril 2025 [en ligne ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/vivant/systemes-symbiotiques-parasites/.
Les articles de l’Encyclopédie de l'environnement sont mis à disposition selon les termes de la licence Creative Commons BY-NC-SA qui autorise la reproduction sous réserve de : citer la source, ne pas en faire une utilisation commerciale, partager des conditions initiales à l’identique, reproduire à chaque réutilisation ou distribution la mention de cette licence Creative Commons BY-NC-SA.


