



The first terrestrial ecosystems
PDF
The establishment of complex continental ecosystems is probably the most important event in the history of multicellular life after the Cambrian and Ordovician radiations that revolutionized the marine world and led to the formation of the first complex ecosystems (see The first complex ecosystems). Plants were the first to colonize the mainland during the Ordovician, 470 million years ago, fundamentally changing the geosphere through oxygenation of the atmosphere, soil formation and the establishment of new climatic and sedimentary regimes. Arthropods were the first animals to take the first steps on land along with myriapods (“centipedes”) and arachnids (spiders, scorpions, mites) at the end of the Silurian, 430 million years ago, then hexapods (insects) followed at the beginning of the Devonian (- 410 million years). Terrestrial vertebrates, tetrapods, only appeared in the middle of the Devonian (-380 million years ago), while the first complex terrestrial ecosystems, formed by highly diversified forest communities, were already well established. However, the anatomy of the first tetrapods reveals that they remained subservient to the aquatic habitats present on the continents (rivers, streams, ponds) in order to integrate literally terrestrial ecosystems only from the Carboniferous (-345 million years).
1. From the first spores to the first forests
Accurately reconstructing the evolutionary history of terrestrial plants and animals is essential to understand the origin and development of terrestrial ecosystems. It also provides an understanding of the biogeochemical cycle of carbon [1] (see A Carbon cycle disrupted by human activities) and its critical impact on the workings of our planet. The carbon cycle is strongly influenced by terrestrial life, mainly through carbon sequestration [2] and atmospheric alteration of rocks. Plants play a very important role in this process, but they are not alone.
MycorrhizaeSymbiotic association between the roots of plants and soil fungi. They affect more than 95% of terrestrial plants. They give plants better access to soil nutrients and help them better resist environmental stresses. facilitate the recovery and assimilation of soil nutrients, thus greatly contributing to rock alteration (see The Biosphere, a major geological player). Arthropods also have a crucial role in soil development and the decomposition and recycling of nutrients.

(i) They are found in terrestrial sediments but their abundance decreases as one moves away from the shoreline.
(ii) Some of them are very similar to liverwort spores, small non-vascularized and thalloid plants (i.e. without morphological differentiation into roots, stems and leaves) which are considered to be the first terrestrial plants.
(iii) Finally, others were preserved in situ during the Lower Devonian in fossils -with exceptional conservation- from bryophytes (the cladeAll or group of organisms whose members, however different they may have become, descend from the same common ancestor group: it is a monophyletic group. In a phylogenetic tree: branch of the tree that contains an ancestor and all his descendants. including liverworts, mosses and hornworts; Figure 2). These bryophytes play a considerable role in rock alteration, and it is possible that they were sufficiently abundant at the end of the Ordovician to trigger a decrease in atmospheric CO2 concentration.
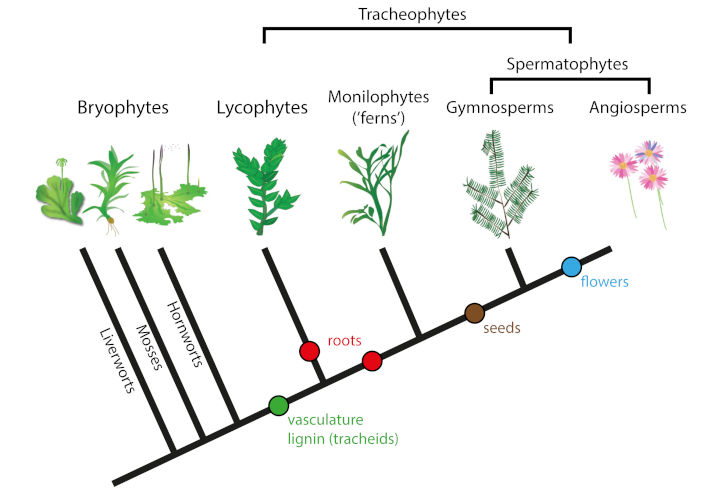
The first fossil remains of stems and/or roots of terrestrial plants come from the Upper Silurian about 430 million years ago. These remains are all small, sometimes minuscule, and often consist of thin carbon films that do not preserve any internal structure. Among these fossils are the first vascular plants, also known as tracheophytes, which have lignified tissues (the tracheids forming the primary xylem) that allow water and mineral salts to be transported through the plant.
Lignin, which fills the spaces of the cell wall, is an important innovation of vascular plants (as well as some algae), providing rigidity, impermeability and resistance to degradation (see The biosphere, a major geological player). Vascular plants diversified between the Upper Silurian and Lower Devonian, but remained small in size, rootless or with a very limited root system, and were therefore confined to moist lowland plains.
The vast majority of our knowledge of early terrestrial plants and ecosystems is due to the outstanding preservation of the Rhynie chert in Scotland. This Lower Devonian deposit (Praguian, 410 million years ago) has produced a wide variety of extraordinarily well preserved terrestrial plants, in particular the oldest traces of rhizoids (fine epidermal growths that help fixing the plant to the ground and absorb water and mineral salts), mycorrhizae and fungal-algae symbiosis (lichens). Wood (secondary xylem), which provides mechanical support for the plant, thus allowing the production and maintenance of trees, also appears in the Lower Devonian.

2. Root systems and soil formation
Tree growth has been accompanied by the development of large root systems. Rooting began in the Praguian with the appearance of very short roots/rhizomes in herbaceous lycophytes, before reaching penetration depths of less than 20 cm in the Eifelian-Givetian and up to nearly one meter in the Frasnian-Famennian with the expansion of Archaeopteridales (Figure 3).The horizontal and vertical increase in root systems strongly affected rock alteration rates and processes, leading to soil formation (or pedogenesis [4]) and landscape stabilization.
Before the Devonian, soils were probably only raw rocks or very thin microbial mats. Paedogenesis was then carried out exclusively by the action of invertebrates forming burrows. However, soil formation at that time was expected to remain limited given the low primary nutrient productivity generated by algal, lichen or bryophyte communities compared to that generated by forest ecosystems. The advent of trees, the expansion of forests and the increase in root penetration during the Middle and Upper Devonian have caused major changes in alteration processes. Indeed, the chemical alteration of soils is mainly due to the different acids released or produced by mycorrhizae, bacterial decomposition and oxidation of organic matter.
Vascular plants, which produce more acids than algae or lichens, and whose much more developed root system offers a larger contact surface, most probably caused an intensification of substrate alteration, thus leading to the formation of soils and their stratification into different horizontal layers called “horizons“; this is the horizonation process (see The biosphere, a major geological player).
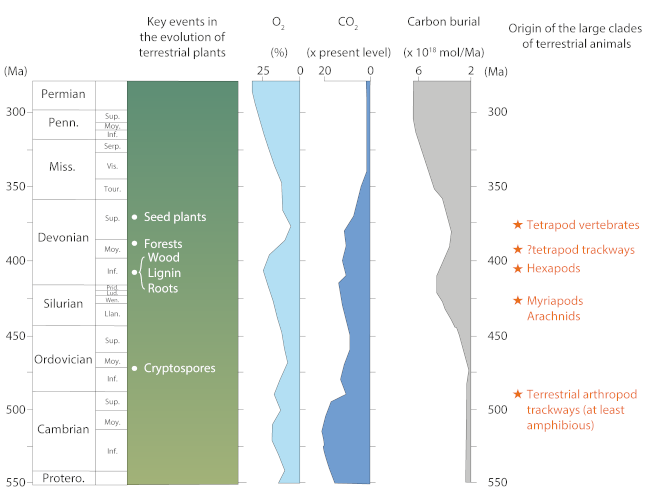
This decrease in CO2 had radical climatic consequences: first, a global cooling of the planet, with a short glacial episode at the very end of the Devonian, and then the establishment of polar ice caps in the southern PangeaSupercontinent formed in the Carboniferous from the collision of existing continents on the Earth’s surface and which then regrouped all the emerged lands. In the Triassic, it split into two continents: Laurasia in the north and Gondwana in the south. during the Carboniferous and Permian (-360 to -260 millions years). Also, the evolution of terrestrial vegetation most likely revolutionized hydrological and sedimentary cycles. The formation of forests increased evapotranspirationProcess resulting from two phenomena: (a) direct evaporation of water from the soil surface to the atmosphere, which is a purely physical phenomenon, and (b) plant transpiration, which is defined as the transfer of water into the plant and the loss of water vapour at the stomata of its leaves. [6] and reduced the albedoReflectivity of a surface, corresponding to the fraction of solar energy that is reflected into space. The more reflective a surface is, the higher its albedo is. [7], promoting the recirculation of atmospheric water andthe increase of precipitation. Also, dense vegetation, due to the high storage capacity of its soil, considerably reduced surface water runoff. Finally, the synergy between land vegetation expansion, reduced runoff and landscape stabilization most probably reduced sediment production.
3. New habitats and landscapes
The presence of root systems developed at the end of the Devonian, ensuring the continuous growth of plants through its nutrient storage and lateral development capacities to find new resources, also allowed the colonization and exploitation of new habitats, hitherto unused or underutilized because they were much more constraining. At the same time, the appearance of the seed offered more possibilities to spermatophytes (seed plants). Seed production “freed” plants them from their dependence on the aqueous medium for reproduction (the encounter of male and female gametes could only take place in water), allowing them to conquer and colonize drier habitats.
The oldest known seeds date back from around 365 million years , at the end of the Famennian. Seed plants diversified at the beginning of the Carboniferous period (-345 million years ago), settling within the ecological niches previously occupied by Archaeopteridales. Due to their similar root systems, the pedogenesis initiated by the latter in the middle of the Devonian was continued in the Carboniferous by the former, but within a wider variety of habitats.
The evolution of vascular plants and the increase in vegetation cover also modified river landscapes (see Alpine alluvial landscapes and biodiversity). While during the Cambrian and Ordovician periods, the river panorama was dominated by rivers flowing over large areas of windblown sand, the development of vascular plants with root systems during the Devonian stabilized the banks and currents, producing meandering or channel rivers and large floodplains.
The strong expansion of forests during the Carboniferous period marks the establishment of narrower, well fixed, and sometimes “braided” channels that may have formed vegetated islands. This expansion also resulted in a significant production of plant debris, particularly the formation of trunk piles that led to the rapid formation of new channels. River systems, on the other hand, also influenced plant evolution with the formation of new biotopes, sometimes very different from each other, thus allowing the colonization of continents by an ever wider range of organisms.
4. Arthropods, the first animal settlers
However, it should not be forgotten that the new ecological niches created during the Devonian by the expansion of terrestrial plants offered much harsher and less stable environmental conditions than marine and river environments. Their colonization therefore required many morphological but also physiological transformations in both plants and animals. The most important innovations for plants were the development of tree structure (lignin, wood), root systems and seed production, thus allowing plants to play a very important role in the biogeochemical cycle of carbon (see above). Animals faced an identical set of challenges: breathing and reproduction out of water, exposure to ultraviolet rays and osmoregulationAll the homeostatic processes involved in regulating the concentration of dissolved salts in the internal fluids and cellular compartments of living beings. Osmoregulation also refers to all the mechanisms of adaptation to the osmotic pressure of the environment surrounding living organisms. [8].
Arthropods were the first animals to venture onto land. They are generally considered “invertebrates” and display a rigid cuticle or shell (the exoskeleton) and a body divided into segments (the metamers), each with a pair of articulated appendages (legs). Today, there are more than 1.2 million species of arthropods, including chelicerates (arachnida and limulidae; more than 110,000 species), myriapods (“centipedes”; more than 10,000 species), crustaceans (more than 65,000 species) and hexapods (collembola or ”silverfish”, and more than one million species of insects). Although it is impossible to estimate the total number of fossil species that have populated the Earth in the past, arthropods have been major components of every ecosystem since the Cambrian, when they started diversifying considerably (see The first complex ecosystems).

Osmoregulation in terrestrial environments has probably been the most difficult challenge faced by animals, but the presence of a waxy cuticle to control water loss offered a huge advantage to arthropods. Similarly, the fact that arthropods have articulated appendages probably greatly facilitated their passage from water to land. Indeed, many aquatic arthropods live on (or in) the substrate, and it seems relatively easy to adapt such a morphology to a terrestrial way of life. Since they can no longer enjoy the buoyancy of the water, terrestrial taxa have developed wider and thicker legs than aquatic ones, which allows them to increase their musculature and thus to be able to overcome the effects of gravity.
Finally, one of the key innovations for arthropods was the development of the tracheal system, an extension of the exoskeleton forming openings on the thorax, the spiracles, opening onto tubes, the tracheas, allowing breathing by simple diffusion of oxygen. This system appeared independently in insects, myriapods and arachnids. Among the latter, scorpions and spiders use “book lungs” (tissue membranes organized like the pages of a book).
The branchiopod crustaceans, the clade that includes triops and artemias, adopted a totally different strategy. More than 365 million years ago, they settled in temporary ponds, small bodies of stagnant fresh water that periodically dry out, in which they thrived thanks to the production of drought-resistant eggs that allowed them -as plant seeds do- to survive dry periods while waiting for the ponds to fill again with water. Although the development and reproduction of these animals takes place in water, their embryonic “egg” stage, which lasts most of their life cycle, is terrestrial, as eggs can remain viable in sediments for up to several years. In the Upper Devonian, other animals remained bound to the aquatic environment, particularly vertebrates.
5. And finally… the vertebrates!
Vertebrates were therefore the last to colonize the continental environments, only at the end of the Devonian. Indeed, the transition to land of vertebrates could not have been as rapid as that of arthropods, better equipped to deal with the numerous anatomical and physiological problems caused by terrestrial life. And unlike arthropods, it seems difficult to believe that many vertebrate groups could have independently left the water for land, given the major anatomical differences between aquatic and terrestrial vertebrates.
During the Devonian, often considered by palaeontologists as the “Age of Fish“, there was an extraordinary diversification of many groups of aquatic vertebrates (commonly known as “fishes“). Some of them still comprises extant representatives, such as:
– chondrichthyans (cartilaginous vertebrates, such as sharks, rays and chimeras, with more than 100 extant species);
– osteichthyans (bony vertebrates, including more than 28,000 extant species).
Others, such as placoderms (armoured vertebrates) and different groups of agnathans (jawless vertebrates), disappeared at the end of this period.
However, it is among osteichthyans that we must look for the aquatic origin of terrestrial vertebrates. Indeed, osteichthyans – which appeared at the end of the Silurian – are divided into two main groups:
– actinopterygians (ray-finned fish), which include the vast majority of “fish” species alive today;
– sarcopterygians (lobe-finned fish), which diversified enormously during the Devonian and some of whose representatives, tetrapods (four-legged vertebrates), were able to leave the aquatic environment a few dozen million years later.
The discovery and study of many fossil sarcopterygian species allows us to better reconstruct the different evolutionary stages of the “fish-tetrapod transition” and to better understand the origin of the characters associated with terrestrial life. One of these first features is the ability to breathe atmospheric oxygen through specialized organs, the lungs. However, their origin is older than the first tetrapods, since it is considered that rudimentary lungs were already present in the first sarcopterygians more than 420 million years ago. Their retention under a vestigial form in the extant coelacanth (the genus Latimeria comprising two species) attest of their long evolutionary history and of their status as a common feature to all sarcopterygians. Indeed, functional lungs associated with the use of gills can still be found in lungfish (or dipnoans) which despite undergoing a very important diversification at the end of the Palaeozoic are currently represented by only six species distributed in three genera (Neoceratodus, Protopterus and Lepidosiren). Thus, the number of sarcopterygian “fishes” is very small in today’s fauna.

The exceptional conservation of its fossils has made it possible to establish the link between the weight-bearing limbs of tetrapods and the fleshy fins of sarcopterygians, since both contain the same bones that form our arms and forearms (Figure 7). The increasingly robust fins and organization of the skull bones and girdles of the most derived tetrapodomorphs, such as the ‘elpistostegalian’ Tiktaalik ([15],[16]) or the recently rediscovered Elpistostege, confirm the surprising idea that the tetrapod features often associated with terrestrial life evolved in aquatic environments and these will only be fully used in terrestrial environments after their transition from water to land.
The oldest known tetrapods date from the late Devonian (Famennian) period. Although other fossil remains (direct or indirect) attributable to tetrapods are older, the famous Ichthyostega and Acanthostega of Greenland are the first to show their most distinctive character: the fingers (Figure 6). Curiously, we noticed that they were polydactile, that is, they bore more than five fingers in each limb (seven for Ichthyostega and eight for Acanthostega!).
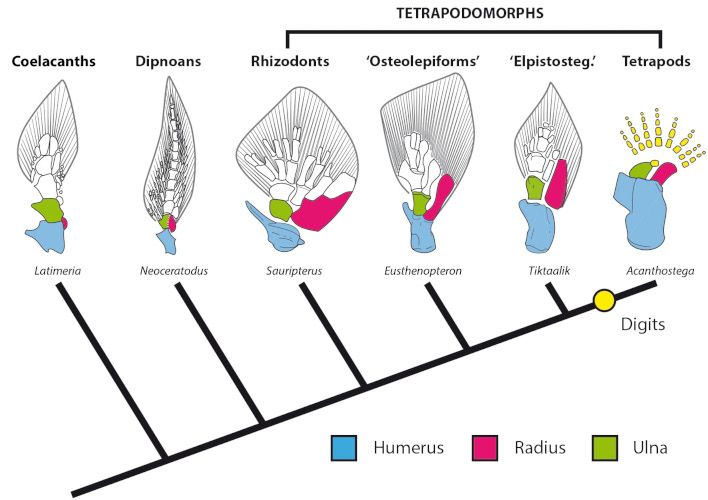
The tetrapods can be thus considered the result of a successful evolutionary experiment towards terrestriality of one of the many groups of tetrapodomorphs fishes populating the coastal or shallow river aquatic environments of the Devonian.
The origin of tetrapods should therefore not be confused with their departure from the water. The origin of tetrapods, which according to recent (but still controversial) discoveries could date back to the Middle Devonian (Eifelian) (see Figure 4), therefore occurred several tens of million of years before the first strictly terrestrial forms appeared. At present, there is no single theory explaining the colonization of the continental environments by vertebrates, but it would seem that the extinction crises of the late Devonian played a crucial role in their abandonment of water.
Thus, at the beginning of the Carboniferous period, many amphibian tetrapod lineages show in parallel an increasingly marked trend towards terrestriality. But it was only in the Upper Carboniferous, about 320 millions years ago, that the appearance of the first amniotesVertebrate taxon grouping together the species in which the embryo and then the foetus are protected by an amniotic sac, called amnios. The young, which develops in a shell or maternal uterus, grows in an aqueous medium, preserved through amnion. This characteristic has allowed these animals to colonize the terrestrial environment and to be permanently removed from the aquatic environment. Amniotes include reptiles, birds and mammals (reptiles sensu lato) and the evolution of the amniotic egg in which the embryo develops in a liquid protected by a series of membranes and an outer shell), allowed vertebrates to complete the transition to land initiated during the Devonian and will contribute to the definitive colonization of all terrestrial ecosystems on the planet by tetrapods now breeding on land. Their impact on the biosphere will be sustainable and will contribute to the stabilization of terrestrial habitat to date.
References and notes
Cover image. Model of reconstruction of Acanthostega at the State Museum of Natural History in Stuttgart (Germany). By Dr. Günter Bechly (Own work) [CC BY-SA 3.0], via Wikimedia Commons.
[1] https://en.wikipedia.org/wiki/Carbon_cycle
[2] https://en.wikipedia.org/wiki/Carbon_sequestration
[3] Algeo T.J. & Scheckler S.E. (1998) Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events. Philosophical Transactions of the Royal Society of London B Biological Sciences 353, 113-130
[4] https://en.wikipedia.org/wiki/Pedogenesis
[5] Gibling M.R. & Davies N.S. (2012) Palaeozoic landscapes shaped by plant evolution. Nature Geoscience 5, 99-105
[6] https://en.wikipedia.org/wiki/Evapotranspiration
[7] https://en.wikipedia.org/wiki/Albedo
[8] https://en.wikipedia.org/wiki/Osmoregulation
[9] https://en.wikipedia.org/wiki/Phylogenetics
[10] Waddington J., Rudkin D.M. & Dunlop J.A. (2015) A new mid-Silurian aquatic scorpion – One step closer to land? Biology Letters 11, 20140815
[11] Engel M.S. & Grimaldi D.A. (2004) New light shed on the oldest insect. Nature 427, 627-630.
[12] https://en.wikipedia.org/wiki/Euthycarcinoidea
[13] http://www.sepaq.com/pq/mig/index.dot
[14] http://www.miguasha.ca/mig-en/the_on-site_museum.php
[15] http://www.devoniantimes.org/
[16] http://tiktaalik.uchicago.edu/
The Encyclopedia of the Environment by the Association des Encyclopédies de l'Environnement et de l'Énergie (www.a3e.fr), contractually linked to the University of Grenoble Alpes and Grenoble INP, and sponsored by the French Academy of Sciences.
To cite this article: GUÉRIAU Pierre, MONDEJAR FERNANDEZ Jorge (January 5, 2025), The first terrestrial ecosystems, Encyclopedia of the Environment, Accessed March 14, 2025 [online ISSN 2555-0950] url : https://www.encyclopedie-environnement.org/en/life/first-terrestrial-ecosystems/.
The articles in the Encyclopedia of the Environment are made available under the terms of the Creative Commons BY-NC-SA license, which authorizes reproduction subject to: citing the source, not making commercial use of them, sharing identical initial conditions, reproducing at each reuse or distribution the mention of this Creative Commons BY-NC-SA license.


