





第一批陆地生态系统

复杂大陆生态系统的建立可能是寒武纪大爆发和奥陶纪大辐射后多细胞生命史上最重要的事件。奥陶纪辐射彻底改变了海洋世界,并形成了第一个复杂生态系统(见《第一批复杂的生态系统》)。4.7亿年前的奥陶纪,植物首先在大陆定居,大气的氧化、土壤的形成以及新的气候和沉积体系的建立,根本上改变了地圈。节肢动物是第一批踏上陆地的动物。4.3亿年前的志留纪晚期,多足类(蜈蚣)和蜘蛛类(蜘蛛、蝎子、螨类)首先登陆,而后在泥盆纪早期(4.1亿年前)六足类(昆虫)登陆。陆地脊椎动物,即四足动物,直到泥盆纪中期(3.8亿年前)才出现,那时高度多样化的森林群落已经形成了第一个复杂的陆地生态系统。然而,第一批四足动物的解剖显示,它们仍然栖息于陆地上的水生栖息地(河流、溪流、池塘),以进一步完善真正意义上的石炭纪(3.45亿年前)陆地生态系统。
1. 从第一个孢子到第一片森林
准确重建陆地动植物的演化史对于理解陆地生态系统的起源和发展至关重要。这也有助于更好地理解碳的生物地球化学循环[1](见《被人类活动干扰的碳循环》)及其对地球运转的关键影响。由于碳固定[2]和岩石的蚀变,碳循环受到陆地生命的强烈影响。植物在这一过程中扮演着非常重要的角色,但它们并不孤单。
菌根加快土壤养分的恢复和吸收,因此极大地促进了岩石的转化(见《生物圈,地质过程的主要参与者》)。节肢动物在土壤发育以及养分的分解和循环中也起着至关重要的作用。

最古老的陆地植物化石可以追溯到近4.7亿年前的中奥陶纪早期。这些化石仅仅是它们的孢子,微化石被保存为孤立的细胞(称为单孢体),或者由两个(二分体)或四个(四分体)联合在一起组成的隐孢子(图1)。下列证据表明这些孢子来自陆地植物:
(1) 它们存在于陆地沉积物中,但离海岸线越远其丰度越低。
(2) 其中一些与苔藓类的孢子非常相似,苔藓是小的非维管束植物和叶状体植物(即没有根、茎、叶分化的植物),被认为是第一种陆生植物。
(3) 最后,在泥盆纪早期,类似的孢子保存在苔藓植物的化石中(该分支包括苔类、藓类和角苔类;图2)。这些苔藓植物在岩石蚀变中起着相当大的作用,很可能正是由于奥陶纪晚期苔藓植物十分丰富,导致大气二氧化碳浓度下降。
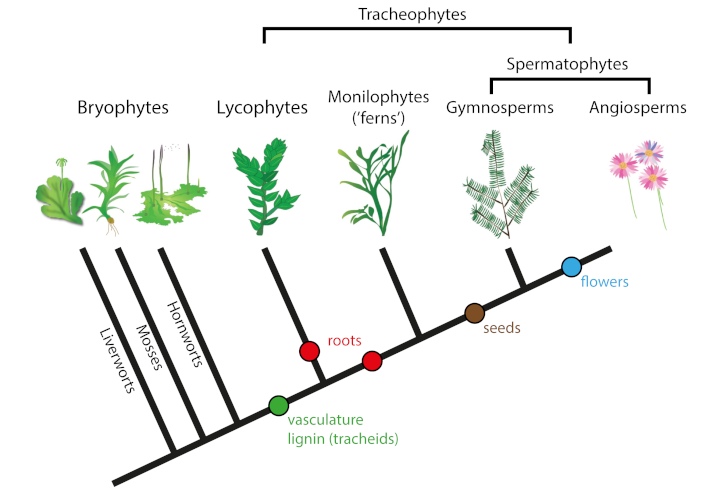
(图2. Tracheophytes 维管植物,Bryophytes 苔藓植物,Lycophytes 石松类,Monilophytes (‘ferns’) 蕨类植物,Gymnosperms 裸子植物,Angiosperms 被子植物,Liverworts 苔类,Mosses 藓类,Hornworts角苔类,vasculature lignin (tracheids) 维管结构木质素(管胞),roots根,seeds 种子,flower 花)
第一批陆生植物茎和/或根的化石来自大约4.3亿年前的志留纪晚期。这些化石都很小,有时是微体的,通常是没有任何内部结构的薄碳膜组成。在这些化石中有最早的维管植物,也被称为导管植物,它们有木质化的组织(形成初生木质部的管胞),以使水和矿物盐通过植物体运输。
木质素填充了细胞壁的空隙,是维管植物(以及一些藻类)的一项重要创新,增强了刚性、不透水性和抗降解性(见《生物圈,地质过程的主要参与者》)。志留纪晚期和泥盆纪早期之间,维管束植物变化多样,但它们体积仍然很小,无根或根系非常有限,因此生长在潮湿的低海拔平原。
我们对早期陆生植物和陆生生态系统的大部分了解都得益于苏格兰莱尼燧石层(Rhynie chert)的完好保存。这个泥盆纪早期的沉积物(布拉格期,4.1亿年前),产生了各种各样保存非常完好的陆生植物,特别是最古老的假根(纤细的表皮生长帮助植物固定在地面上,吸收水分和矿物盐)、菌根和真菌-藻类共生(地衣)。木材(即次生木质部)也出现在泥盆纪早期,能够为植物提供机械支持,使得树木能够生产和维持。
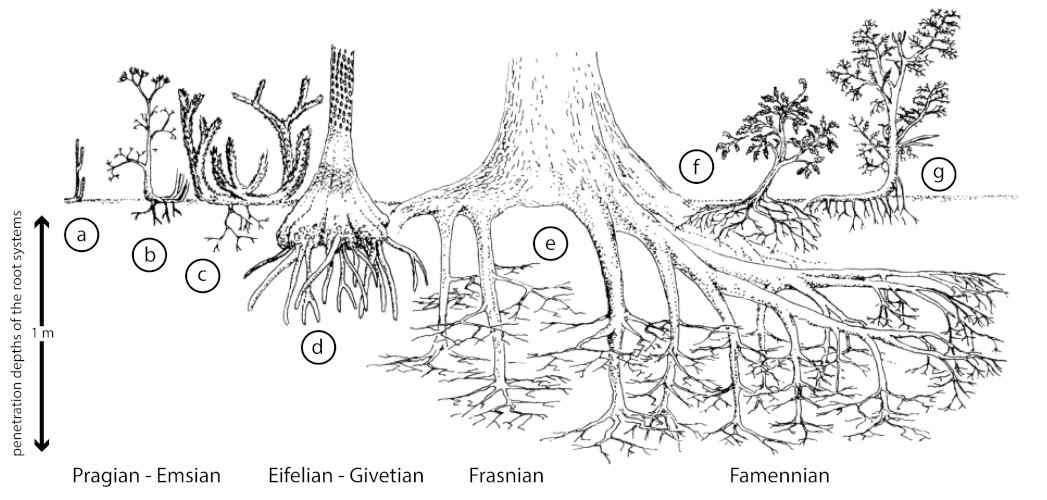
2. 根系与土壤形成
树木的生长伴随着大型根系的发育。植物生根始于早泥盆世布拉格期的草本石松类,根/根茎非常短,然后在艾菲尔阶-吉维特阶,达到近20厘米的穿透深度,在弗拉斯阶-法门阶,随着古羊齿类的扩张而达到近一米的穿透深度(图3)。根系的水平和垂直扩张强烈影响了岩石转化的速率和过程,从而形成土壤(成土作用[4]),景观也得以稳定。
在泥盆纪之前,土壤可能只是原始岩石或非常薄的微生物垫。那时,土壤的形成由形成洞穴的无脊椎动物来完成的。然而,考虑到藻类、地衣或苔藓群落产生的初级养分生产力低于森林生态系统,这一时期的土壤形成仍然受到限制。在泥盆纪中期和晚期,树木的出现、森林的扩张和根系渗透率的增加,带来了蚀变过程的重大变化。事实上,土壤的化学变化主要是由菌根释放或产生的各种酸、细菌分解和有机物氧化引起的。
维管植物产生的酸比藻类或地衣更多,其发达的根系提供了更大的接触面,很可能加剧了基底变化,从而导致土壤的形成,并将其分层为不同的水平层,称为“土壤层”;这就是分层作用的过程(见《生物圈,地质过程的主要参与者》)。
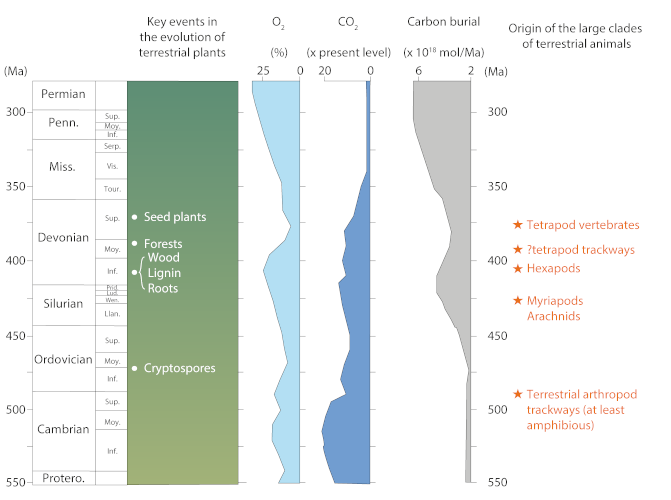
(Permian 二叠纪,Penn. 宾夕法尼亚纪,Miss. 密西西比纪,Devonian 泥盆纪,Silurian 志留纪,Ordovician奥陶纪,Cambrian寒武纪,Protero. 原生代,Sup. 上,Moy. 中,Inf. 下,Serp. 谢尔普霍夫阶,Vis. 维宪阶,Tour. 杜内阶,Prid. 普里道利统,Lud. 罗德洛统,Wen. 温洛克统,Lian. 兰多维列统,Seed plants 种子植物,Forests 森林,Wood 木材,Ligin 木质素,Roots 根,Cryptospores 隐孢子,Key events in the evolution of terrestrial plants 陆生植物进化过程中的关键事件,Carbon burial碳埋藏,Origin of the large clades of terrestrial animals 陆地动物大分支的起源,Tetrapod vertebrates 四足脊椎动物,Tetrapod trackways 四足动物足迹,Hexapods 六足动物,Myriapods 多足,Arachnids蛛形纲动物,Terrestrial arthropod trackways (at least amphibious) 陆生节肢动物的足迹(至少两栖))
泥盆纪晚期大面积森林的建立,产生了大量光合作用活动,导致了大气中强烈的固碳和氧化作用(图4,[5])。随着岩石蚀变和土壤形成增强,其对碳的捕获作用也增强,碳的整体生物地球化学循环被破坏,大气中CO2浓度迅速下降。
二氧化碳的减少导致气候急剧变化:首先是全球变冷,泥盆纪末期有一段短暂的冰期,然后在石炭纪和二叠纪(-3.6亿到-2.6亿年)期间,盘古山脉南部形成了极地冰盖。此外,陆地植被的演变很可能彻底改变了水文和沉积循环。森林的形成导致蒸散量增加[6],反照率降低[7],促进了大气水分再循环和降水。此外,茂密植被下的土壤蓄水能力高,地表水径流大大减少。最后,陆地植被扩张、径流减少和景观稳定三者很可能协同减少了沉积物的产生。
3. 新的栖息地和景观
根系形成于泥盆纪末期,通过养分储存和寻找新资源的横向发展能力,支持植物持续生长,也能够扩张到新栖息地。到目前为止,部分栖息地之所以没有被利用或没有得到充分利用,是因为植物受到了更多的限制。种子的出现为种子植物提供了更多的可能性,使它们的繁殖摆脱了对水介质的依赖(雄性和雌性配子的相遇只能在水中发生),也能够征服更干燥的栖息地并定居。
已知最古老的种子可以追溯到大约3.65亿年前,也就是泥盆纪法门期末期。在石炭纪初期(-3.45亿年前),种子植物繁盛于以前由古羊齿类占据的生态位中。古羊齿类于泥盆纪中期引发了成壤作用,而后在石炭纪由种子植物继续完成,但覆盖的生境范围更加广泛。
维管植物的进化和植被覆盖率的增加也改变了河流景观(见《高山冲积景观和生物多样性》)。在寒武纪和奥陶纪,河流全貌以流经大片风积沙为主,而泥盆纪期间,维管植物的根系发育稳定了河岸和水流,形成了曲流或河道,以及大片河漫滩。
石炭纪期间森林的强劲扩张标志着形成更狭窄的、更固定的、有时甚至是“交织”的河道,也许正是这些河道造就了植被繁茂的岛屿。森林扩张还产生了大量植物残渣,特别是树干丛,也推动了新河道迅速形成。另一方面,河流系统影响了植物的进化,形成了新的群落生境,有时这些生境间差异较大,从而使得越来越多的生物能在大陆上定居。
4. 节肢动物,第一批动物定居者
然而,我们不该忽视,这些泥盆纪期间陆生植物扩张所形成的新生态位,环境条件比海洋和河流环境更恶劣、更不稳定。因此,植物和动物需要进行许多形态和生理上的转变才能定居。对植物来说,最重要的创新是树木结构(木质素、木材)、根系的发展和种子的产生,使植物在碳的生物地球化学循环中发挥非常重要的作用(见上文)。动物面临着一系列相同的挑战:离开水中进行呼吸和繁殖,暴露在紫外线下,以及渗透调节[8]。
节肢动物是第一批登上陆地的动物。它们通常被认为是“无脊椎动物”,有一个坚硬的角质层或外壳(外骨骼)和一个被分成几个部分的身体(体节),每个部分都有一对关节附肢(腿)。目前节肢动物有120多万种,包括螯肢类(蜘蛛纲和蝇科;11万多种)、多足类(“蜈蚣”;10000多种)、甲壳类(6.5万多种)和六足类(弹尾目或“蠹虫”,以及100多万种昆虫)。虽然无从估计过去地球上化石物种的总数,但自寒武纪以来,节肢动物一直是每个生态系统的主要组成部分。正是那时,节肢动物变得越来越多样化(见《第一批复杂的生态系统》)。

现存的陆地节肢动物在土壤发育以及养分的分解和循环中起着至关重要的作用。它们的亲缘关系(系统发育[9])表明,至少有七类节肢动物独立进入了大陆:多足类、蜘蛛类、六足类,以及四类甲壳类动物(包括林虱和螃蟹)。最古老的陆地节肢动物化石可以追溯到志留纪,由多足类和蜘蛛类(螨类、蜘蛛类和蝎子类)组成(图4、图5,[10], [11])。但在寒武纪海岸沉积物中发现的“脚印”(足迹),很可能是直虾纲动物留下的[12],它们是一种已灭绝的神秘节肢动物,这有力地表明陆生节肢动物早在志留纪之前就存在。与植物一样,莱尼燧石层(Rhynie chert)也是许多奇妙发现的发源地。在其中发现了最古老的六足动物化石,可以追溯到泥盆纪早期,包括弹尾目、“蠹虫”(Leverhulmia maria)和最古老的昆虫:莱尼虫(Rhyniognatha hirsti)的下颌。
陆地环境中的渗透调节可能是动物面临的最大挑战,但蜡质角质层可以控制水分流失,这是节肢动物很大的优势。同样,节肢动物的关节附肢可能极大地推动它们从水中登上陆地。事实上,许多水生节肢动物生活在水体基底上(或基底中),以这种形态适应陆地的生活方式似乎相对容易。由于不能再利用水的浮力,陆生类群的腿比水生类群更宽更粗,让它们能够增加肌肉,从而克服重力的影响。
最后,节肢动物的一项重大进步是发展了气管系统,外骨骼在胸腔上形成开口的延伸,气孔通向气管,通过简单的氧气扩散进行呼吸。该系统在昆虫、多足类和蜘蛛纲动物中独立出现。其中,蝎子和蜘蛛使用“书肺”(像书页一样组织起来的组织膜)。
鳃足类甲壳动物(包括鲎虫和卤虫)采取了完全不同的策略。3.65亿多年前,它们在临时池塘里定居。临时池塘是一小片死水,会定期干涸。但抗旱的卵让它们得以茁壮成长,像植物种子一样,在干旱期存活下来,等待池塘再次注入水。虽然这些动物的发育和繁殖是在水中进行的,但它们胚胎发育的“卵”(它们一生大部分时间的形态)阶段是陆生的,因为卵可以在沉积物中存活长达数年。在泥盆纪晚期,其它动物仍然离不开水,特别是脊椎动物。
5. 最后是……脊椎动物!
脊椎动物直到泥盆纪末期才在大陆定居的,是定居的最后一类动物。事实上,脊椎动物向陆地的过渡不可能像节肢动物那样迅速,节肢动物更有能力应对在陆地生活的众多解剖学和生理学难题。与其不同,水生和陆生脊椎动物之间存在较大解剖学差异,似乎很难相信许多脊椎动物群体可以独立离水登陆。
古生物学家通常认为泥盆纪是鱼的时代,在泥盆纪,许多水生脊椎动物(俗称“鱼”)展现出非同寻常的多样性。其中一些代表至今依然存在,例如:
–软骨鱼类(软骨性脊椎动物,如鲨鱼、蝠鲼和银鲛,现存物种超100种);
–硬骨鱼类(硬骨脊椎动物,包括28,000多个现存物种)。
其他动物,比如盾皮鱼类(盔甲脊椎动物)和不同种类的无颌类(无颌脊椎动物),在这一时期结束后再无影踪。
然而,我们必须在硬骨鱼中寻找陆生脊椎动物的水生起源。出现在志留纪末期的硬骨鱼主要分为两类:
–辐鳍鱼类(辐鳍鱼),涵盖了现存的绝大多数“鱼”;
–肉鳍鱼类(肉鳍鱼),在泥盆纪发生了巨大的变化,其中的代表四足类(四足脊椎动物)在几千万年后已经可以离开水环境。
许多肉鳍鱼类化石的发现和研究使我们能够更好重建“鱼类-四足动物转变”的不同进化阶段,并能更好地理解与陆地生命相关特征的起源。这些最早的特征之一是能够通过特殊的器官——肺来呼吸。然而,它们的起源比最初的四足动物更古老,因为人们认为在4.2亿多年前,第一批肉鳍鱼类就已经有了原始的肺。这些肺在现存的腔棘鱼(Latimeria属,由两个物种组成)中以残留的形式保留下来,证明了它们漫长的进化历史,以及它们作为所有肉鳍鱼类的共同特征的地位。事实上,与鳃相关功能的肺仍然可以在肺鱼(或双鳍金枪鱼)中找到,虽然它在古生代末曾非常多样,但目前仅存三个属(新角鱼属(Neoceratodus))、非洲肺鱼属(Protopterus)和南美肺鱼属(Lepidosiren))的六个物种。因此,在今天的动物类群中,肉鳍鱼类“鱼”的数量非常少。

[图源:(a) © 马蒂厄·杜普伊(Mathieu Dupuis), 加拿大米加沙国家公园(Miguasha National Park); (b and g) © 劳尔·马丁(Raúl Martín);(c) © 约翰娜·科尔(Johanne Kerr), 加拿大米加沙国家公园;(d-f) ©詹妮弗·克拉克(Jennifer Clack)]
然而,这些鱼和四足动物之间的形态差异太大,难以准确地重建“鱼-四足动物转变”的进化史。为了更好地了解四足动物的起源,需要参考四足形目(肉鳍鱼类,与四足动物的亲缘关系比肺鱼更近)和它们的许多典型化石。在泥盆纪和石炭纪,四足动物在海岸和淡水环境中也表现出了多样性,在那里它们是捕食者,通常体型很大(“骨鳞鱼目”的伊甸鳍鱼(Edenopteron)或超过两米,而根齿类的根齿鱼(Rhizodus)可能长达六米!)。魁北克最著名的物种之一是魁北克上泥盆纪(弗拉斯阶)“骨鳞鱼目”真掌鳍鱼(Eusthenopteron)(图6)[13], [14]。
其化石保存地如此完好,让人们能够在四足动物的负重四肢和肉鳍鱼类的肉鳍之间建立联系,因为它们具有构成我们手臂和前臂的相同的骨骼(图7)。衍生最多的四足形目动物,如“希望螈类”提塔利克鱼(Tiktaalik) [15], [16]或最近发现的希望螈(Elpistostege),越来越结实的鳍、颅骨和腰带的组织证实了一个令人惊讶的观点,即四足动物的特征通常与在水生环境中进化的陆地生物有关,当它们从水过渡到陆地之后,这些特征才能在陆地环境中得到充分利用。
已知最古老的四足类可以追溯到泥盆纪晚期(法门纪)。虽然其他(直接或间接地)归于四足动物的化石遗迹更古老,但格陵兰岛著名的鱼石螈(Ichthyostega)和棘鱼石螈(Acanthostega)最先显示出它们最独特的特征:手指(图6)。奇怪的是,我们注意到它们是多指的,也就是说,它们的每条腿上都有五个以上的手指(鱼骨有七个手指,棘骨有八个手指!)。
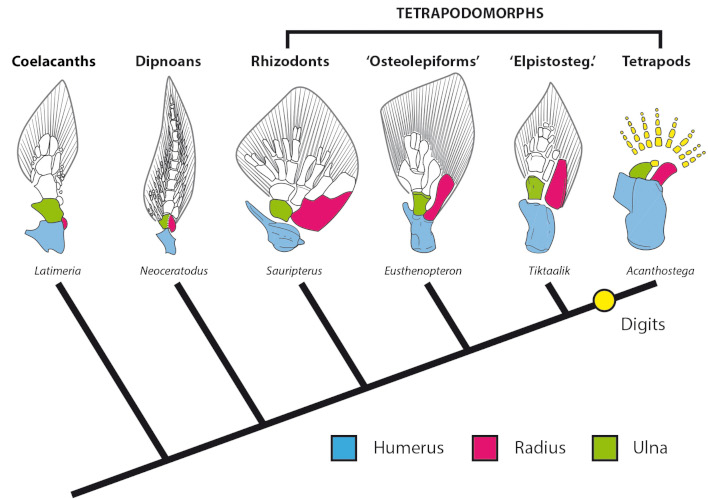
然而,尽管这些早期的四足动物有真实的四肢,即由三部分组成的骨骼附肢:顶足类(手臂)、斑足类(前臂)和自肢(手),这是陆生脊椎动物的特征(图7),但它们显然是水生动物。对它们的解剖学研究阐明了与水中生命密切相关的特征的保持,如内鳃的存在,发育良好的尾鳍上的射线,以及横跨头骨不同部位并沿着躯干延伸的感觉侧线。
因此,四足动物可以被认为是在泥盆纪沿海或浅水河流水环境中,众多四足形鱼中的一种向陆地成功进化的结果。
因此,四足动物的起源不应与它们离开水面混为一谈。根据最近的发现(仍有争议),四足动物的起源可以追溯到泥盆纪中期(艾菲尔阶)(见图4),因此,四足动物的起源发生在第一批严格意义上的陆地种类出现的几千万年前。目前,还没有单一的理论来解释脊椎动物在大陆环境的定居,但似乎泥盆纪晚期的物种灭绝危机在它们弃水的过程中起到了至关重要的作用。
因此,石炭纪初期,许多两栖四足动物谱系同时显示出日益明显的陆相趋势。但直到大约3.2亿年前的石炭纪早期,第一批羊膜卵动物(爬行动物Sensu Lato)的出现和羊膜卵的进化(胚胎在一系列膜和外壳的保护下在液体中发育),才使脊椎动物完成了泥盆纪开始的陆地过渡,并有助于目前在陆地上繁殖的四足动物最终定居在地球上的所有陆地生态系统中。它们对生物圈的影响将是可持续的,并将有助于迄今为止陆地栖息地的稳定。
参考资料及说明
封面图片:德国斯图加特国立自然历史博物馆的棘鱼石螈(Acanthostega)重建模型。作者:君特·贝赫利博士(Dr. Günter Bechly)((原创)[CC BY-SA 3.0协议],维基共享资源。
[1] https://en.wikipedia.org/wiki/Carbon_cycle
[2] https://en.wikipedia.org/wiki/Carbon_sequestration
[3] Algeo T.J. & Scheckler S.E. (1998) Terrestrial-marine teleconnections in the Devonian: links between the evolution of land plants, weathering processes, and marine anoxic events. Philosophical Transactions of the Royal Society of London B Biological Sciences 353, 113-130
[4] https://en.wikipedia.org/wiki/Pedogenesis
[5] Gibling M.R. & Davies N.S. (2012) Palaeozoic landscapes shaped by plant evolution. Nature Geoscience 5, 99-105
[6] https://en.wikipedia.org/wiki/Evapotranspiration
[7] https://en.wikipedia.org/wiki/Albedo
[8] https://en.wikipedia.org/wiki/Osmoregulation
[9] https://en.wikipedia.org/wiki/Phylogenetics
[10] Waddington J., Rudkin D.M. & Dunlop J.A. (2015) A new mid-Silurian aquatic scorpion – One step closer to land? Biology Letters 11, 20140815
[11] Engel M.S. & Grimaldi D.A. (2004) New light shed on the oldest insect. Nature 427, 627-630.
[12] https://en.wikipedia.org/wiki/Euthycarcinoidea
[13] http://www.sepaq.com/pq/mig/index.dot
[14] http://www.miguasha.ca/mig-en/the_on-site_museum.php
[15] http://www.devoniantimes.org/
[16] http://tiktaalik.uchicago.edu/
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: GUÉRIAU Pierre, MONDEJAR FERNANDEZ Jorge (2024年12月1日), 第一批陆地生态系统, 环境百科全书,咨询于 2025年1月21日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/first-terrestrial-ecosystems-2/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。


