





平衡在稳定性和变异性之间的基因组

由于身体内外意外的错误或因子影响,生物体的基因组会永久地受到影响。幸运的是,酶纠正了其中大部分异常,从而加强了遗传的稳定性。但酶也会导致变异。同时,其他系统可以产生新的基因组合。这些突变和重组对于种群适应环境变化和进化至关重要。
1. 引言
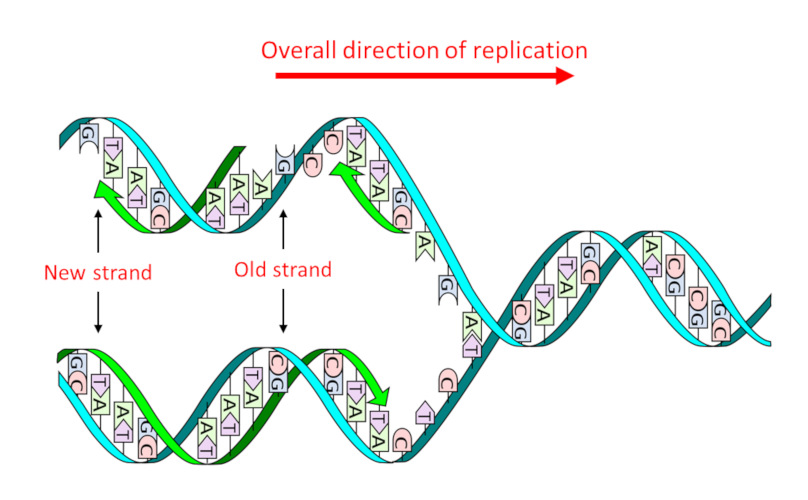
DNA由双序列的单体分子单体分子:分子结构,可通过与相同或不同分子的连续序列,形成聚合物分子结构。(核苷酸)组成,单体分子有四种类型,根据其含氮碱基或核碱基核碱基:在核酸中以核苷酸的形式存在的有机氮化合物,在RNA中与糖结合,在RNA中与核糖结合,在DNA中与脱氧核糖结合。在遗传学上,它们通常被简称为核酸的碱基。可分为:腺嘌呤(A)、鸟嘌呤(G)、胸腺嘧啶(T)和胞嘧啶(C)。这四种类型的碱基精确排序,也就是它们的精确序列,构成了遗传信息的“文本”。其结构为两条互补的链,其中A总是面对T,G总是面对C,即著名的双螺旋结构(图1),该结构有两个主要优势:
- 它允许,通过一个单一的步骤,复制复制:获得两个与初始分子相同的分子的过程。形成两个相同的子代分子(图1),从而通过细胞分裂和有性繁殖传递遗传信息。
- 它可以修复出现在其中一条链上的损伤,同时创造重组分子,即新的遗传组合。因此,这种双链结构提升了基因组的稳定性和变异性。
DNA的遗传变异主要分为两大类:突变和重组。
- 突变是活细胞或病毒基因组的突然变化。在多细胞生物中,如果突变影响了提供配子的生殖细胞,它们就会传递给后代,而且会因此成为可遗传变异。
- 重组是指通过现有遗传物质的交换产生新的遗传组合。重组类型有多种,其发生的生物学机制和作用各异。传统意义上的重组定义只涉及同一物种的DNA。我们现在知道,不同物种之间有自发的基因交换,这被称为转基因,我们也将其纳入这一类别。
这两大变化类型本身又可继续细分。我们将努力提取最相关的要素,以澄清它们各自在种群动态和进化中发挥的作用。
2. 基因突变
2.1. 一般信息
DNA编码的遗传信息的稳定性可能会受到复制酶(执行复制的酶)出错的影响。如果出错,这种酶会在新链中某个含氮碱基的位置放入另一个碱基。但除了复制,DNA也会永久受损。区分损伤和突变很重要(参见遗传多态性和变异)。损伤是DNA物理结构发生异常,在大多数情况下,这会阻止DNA的复制。因此损伤不会遗传给后代。相对而言,突变的分子物理结构正常。只有碱基的序列以及其所含信息遭到修改。因此,它可以正常复制,并将突变传递给子代细胞。
损伤多种多样,涉及到大量的修复酶,每种修复酶都专门针对一种特定类型的损伤。这又称为DNA修复的“细胞工具箱”(该研究成果被授予2015年诺贝尔化学奖)。矛盾的是,正是这些修复系统无意中造成了原发性损伤的实际突变。这些损伤可能由体内(内源性)或外部环境(外源性)的不同因子引发。下面我们来举例说明这些概念。
- 内源性因子:据估计,在温血动物中,由于分子搅动,每个细胞内的DNA分子每天可经历2万至4万次单链切割单链切割:,切割发生在核酸片段链上两个相邻核苷酸之间。。也可能存在碱基的丢失。据估计,每个细胞每天损失10000个T和10000个C和500个A和500个G。在这些事件中,我们甚至可以将其称之为自发性损伤。最常见的内源性因子是氧衍生物(自由基或活性氧ROS: Reactive oxygen species,“活性氧物种”或“活性氧物种”的缩写。由氧产生的自由基,非常活泼,毒性很大。经常使用缩写ROS,在法语中也常常这样使用。),它是呼吸代谢的正常副产物,在氧化核碱基方面起着重要作用。此外,还有转座元件转座元件: DNA序列,有时称为转座子,通过一种叫做转座子的机制,能够在基因组中自主移动。这些可移动的DNA序列是分散重复序列的一部分,被认为是进化和生物多样性的强大驱动力。,我们将在下文进一步讨论。
- 外源性因子:它们可以是物理(辐射)或化学因素。最常见的是紫外线(UV),其影响通常仅限于皮肤,因为其穿透力不是很强(参见太阳紫外线的细胞影响)。例如,据估计,日光浴会让每个细胞在每小时内,同一条DNA链的相邻胸腺之间产生6万到8万个异常化学键,每一个异常化学键都能阻止复制。如果没有全部修复,细胞就会死亡:即皮肤晒伤。
因此,很明显,DNA的稳定性是一个动态过程,是产生损伤和修复损伤永久平衡的结果。这些修复机制不会以恒定水平运行,而是会受到调节。
第一种类型的调节取决于细胞内损伤的数量。这一现象最早出现在大肠杆菌(大肠埃希菌)中,大肠杆菌是遗传学家首选的一个研究对象。早在1974年,人们就认为存在一种称为SOS[2]的响应,它根据损伤的数量调节干预多个修复系统。出现少量损伤时,这种响应增加了忠实修复机制的有效性。但超过一定的损伤阈值,这些机制就不堪重负了。这种情况下,SOS响应诱导合成一种复制酶,这种酶能够修复特定损伤(见下文),但存在一定的误差率。这被称为SOS突变。在20世纪90年代末,研究表明这种突变有助于细菌种群适应恶劣环境,但代价是有害突变会造成重大损失。这是最后一次机会,因此得名为SOS。

根据生物的类型,第二种调节关系到复制的保真度。在所有活细胞、细菌和其他细胞中,这种复制除了DNA复制本身以外,还涉及一种纠正酶系统。由于旧链,该系统纠正了复制酶在新链上所造成的错误,因此错误率非常低,约为百亿分之一(10-10)。这样的准确率对于拥有DNA大分子的生物体来说至关重要:大肠杆菌中有400万个碱基对,人类基因组有30亿个碱基对。
但在多细胞生物中,细胞数量可能非常多:在人类物种中,估计有100万亿(1014)个细胞。在这些生物体中,大多数突变是中性突变,因为基因组包含大量非编码序列(参见遗传多态性和变异)。即使考虑到这一点,100亿分之一(10-10)的错误率仍然太高。此外,由于许多突变也发生在复制之外的其他过程,这就更加明显了。如果每个细胞只复制一个基因,其中许多复制将携带有害突变,有机体将无法存活。因此,对这些生物体来说,有两组染色体(即二倍体,二倍体:其特性为每个细胞的染色体成对存在 (2n条染色体)。这一概念通常与单倍体形成对比,单倍体是指拥有单个复制染色体(n个染色体)的细胞。当生物体的细胞是二倍体时,也就称该生物体或生物体的一部分是二倍体。)是有利且必要的。大多数使一个基因失活的突变是隐性突变,所以一次功能性复制就能使细胞正常工作。简而言之,二倍体就像一个备用轮。
如果我们现在来看病毒的例子,情况就非常不同了,因为大多数病毒的基因组都非常小。对于DNA病毒,一般没有与复制酶相关的纠正活性,因此突变率要高得多,突变率根据病毒种类从10-8到10-6不等。对于基因组由RNA组成的病毒来说更是如此,因为RNA复制酶比DNA复制酶准确率低很多,错误率是10-6到10-4。
然而,可以根据基因组的大小将这些RNA病毒分为两类。在基因组大小在10000个核苷酸左右的病毒中,复制的保真度仅取决于复制酶。流感病毒(13500个核苷酸)和导致艾滋病的HIV病毒(10000个核苷酸)就是这样的例子。在后一种病毒中,每个基因组和每个复制周期的突变率估计为每10个基因组1个点突变。由于每个受感染细胞每天产生的病毒颗粒(病毒粒子)约为10000个,可以看出,受感染宿主的总体种群具有相当大的突变性。
在基因组约有3万个或更多核苷酸的RNA病毒中,现在已知其复制酶具有纠正活性。冠状病毒就是这样,包括导致COVID-19大流行的SARS-CoV-2病毒,其基因组含有29903个核苷酸。这种纠正活性使复制保真度提高了20倍,从而避免产生过多的缺陷病毒粒子。
更笼统地说,在所有病毒中,无论是DNA还是RNA,突变率和基因组大小之间似乎确实存在负相关关系。每一种病毒经过自然选择后,突变率已经达到平衡。突变率不应该太高,这样就不会影响病毒种类的延续,但它仍然足以让病毒变异,从而绕过宿主的防御机制。在这最后一点上,艾滋病病毒就很有代表性。
因此,我们就突变在进化过程中的作用可以发表两点看法:
- 突变在物种适应环境方面发挥着关键作用,通过增加遗传多样性,突变在自然选择的过程中可以发挥作用。我们前面讨论的病毒就是一个例子。耐抗生素细菌、耐杀虫剂昆虫或耐除草剂植物的出现则提供了更多示例。
- 基因突变并非天生有利或不利,而是取决于环境。例如,在亚南极带的克尔格伦群岛上,有一种无翅苍蝇(Calycopteryxmoseley)(图3)。这种特性在我们生活的地区是非常不利的,但在克尔格伦群岛上却是有益的,因为这些苍蝇就不至于被常年肆虐克尔格伦群岛的狂风卷进海洋。

2.2. 点突变
这可能是由于复制过程中的错误,或者更常见的是由于修复不良损伤;点突变产生相同基因(等位基因)的变异。在大肠杆菌中,是SOS系统导致了紫外线照射后的绝大多数突变。上面提到的大多数例子主要与点突变有关。
2.3. 染色体变异
染色体突变是由包括辐射(紫外线除外)在内的导致DNA断裂的因素引起的重排。几个双链断裂可能导致或多或少的重大重排,这取决于它们的数量和涉及片段的大小。
如果两个相距较远的双链断裂(Double-stranded breaks, DSB)发生在同一条染色体上,断裂点之间的整个DNA区段可以有如下后果:(a)丢失,导致个体死亡,(b)在同一位点倒转(inversion倒位),或(c)转移到另一条发生断裂的染色体上(translocation易位)。倒位和易位在自然种群中相当频繁。这些染色体突变干扰减数分裂减数分裂:真核生物中的细胞双重分裂过程,发生在生殖系细胞(二倍体)中以形成配子(单倍体)或生殖细胞。中的染色体配对,从而导致不育。因此,它们可以参与物种形成物种形成:导致新的活物种出现的进化过程,,这些新物种从属于原始物种的种群中分离出来。过程。通常情况下,相邻物种的染色体重排存在差异。
2.4. 染色体组突变
这类突变的机制不同于与前面几类。这些异常不是由原发性损伤引发的,而是由细胞分裂过程(有丝分裂有丝分裂: 是指细胞分裂的染色体事件,是真核细胞周期的一个阶段。这是母细胞染色体的非有性/无性复制(不同于减数分裂)的步骤以及它们在两个子细胞中的平均分配。或减数分裂)中的机械错误引发的。有些可能导致个体出现两个以上的染色体组(2N),但总是整数倍的染色体组(3N, 4N…),这就是多倍体(参见聚焦多倍体)。它们可以存活,因为“基因平衡”得到了尊重:所有基因都有相同的复制数。另一方面,三倍体是不育的,因为配子的遗传平衡必然异常。减数分裂显然不能均匀地在配子中分配奇数倍染色体。多倍体4N、6N或8N是可育的。但它们立即创造了一个新物种,因为任何与二倍体亲代杂交的后代都将不育,这些后代的减数分裂非常不平衡。
减数分裂的其他“失败”可能导致某些配子的染色体不平衡,因此会在后代体内发现。例如,它们将是2N-1(单体)或2N+1(三体),也就是我们之后会谈到的非整倍体,这是一种有害的情况,因为遗传平衡出现问题。
3. 重组
自然界存在许多具有非常不同的生物学作用的重组形式。
3.1. 同源重组

同源重组是已知最常见也是最古老的重组类型。顾名思义,同源重组是在相同的DNA分子之间进行的(图4)。在减数分裂期间,二倍体生物的同源染色体之间经常发生交叉。这是一种“软”变异性:重组在完全相同的位点进行,因此染色体上的基因排列不会发生突变或修改。这导致新的可存活的遗传组合,在给定环境中,可能有不同于亲代组合的适应能力。在细菌中,它也可以发生在接合[6]过程中,从而允许细胞之间进行遗传交换[7]。
这种重组机制也参与了DNA的修复。在二倍体生物中,由辐射引起的染色体双链断裂可以通过在相应片段中的完整同源分子来修复。
3.2. 其他类型的重组
为简化起见,我们将生物学作用迥异的各种事件归类。它们可归为插入重组,因为在大多数情况下,它们会导致基因组中增加DNA片段。
其中最常见的机制是移动遗传元件(也称为转座元件)[8],只含有几百或数千个核苷酸,至少具有“跳跃”到宿主基因组所需的功能。它们存在于细菌及最复杂的真核生物中,有时数量庞大。前者在接合过程中可以在不同物种之间进行交换[6]。由于它们通常携带抗生素耐药性基因,便成为这些耐药性在致病菌中迅速传播的主要因素之一,从而导致严重的医学问题。
在人类物种中,虽然编码所有蛋白质的基因本身只占细胞核DNA的2%至3%,但各种转座元件家族却占了近50%。其中大多数在远古地质时代就已经在我们的谱系中建立起来了,远远早于人类物种的出现。逆转录病毒逆转录病毒:家族RNA病毒。有一种酶,逆转录酶,它允许病毒RNA转录成一种特定的DNA分子,能够与宿主细胞的DNA结合。(HIV属于该家族)既是病毒又是转座元件,仅此一项就占我们DNA的8%。幸运的是,对于保持基因组稳定性来说,这些元件只有小部分仍可移动。与“经典”突变一样,如果在基因中插入转座元件可能有害,但这也可以带来有意义的基因创新,比如说改变基因(表达)调控,或是改变基因自身功能。多项数据表明,它们对基因组和物种的进化有贡献[9]。
最后,有必要提到转基因现象,即不同物种之间的遗传交换。自上世纪90年代末以来,这种现象似乎比我们想象的要频繁得多。之前认为物种屏障在自然条件下不可逾越,但实际情况是,至少在进化的尺度上,物种屏障相当脆弱。这也是创新的来源,成就了基因组和进化的可塑性。
4. 结论
我们简要介绍了不同类型的基因组变异。我们无法详细介绍修复和重组所涉及的酶系统,单是这些就需要写一篇完整的文章(即便不需要写两篇文章!)对这些机制感兴趣的读者可以参考以下网站[10]。最后,有两点需要强调:
- 遗传变异随机发生。这与拉马克进化论的假设不一样,遗传变异不会受环境的引导而去适应环境。已经有大量实验对此加以证明(参见拉马克和达尔文:两种对待生命的不同看法)。然而,与20世纪80年代以前人们的想法相反,其频率会受到环境的调节。我们已经在细菌、果蝇等生物中看到了这一点,细菌中SOS反应使得突变率上升,而在果蝇和拟南芥中,辐射激活同源重组。这些并不是唯一能够调节频率变化的因素,许多生理压力都可以导致这个结果。例如,在细菌培养中使用抗生素也会引发SOS突变,从而增加突变的频率[11,12]。《适应:应对环境挑战》一文中也涉及到这一主题。
- 本文核心论述了生物稳定性/可变性的辩证统一。两者先天对立,但实际上又相辅相成。两者对于种群生存和物种进化都至关重要。
稳定性是指当环境相对稳定时,种群能够持续地适应环境;而变异性是指当环境发生变化时,它能够促进遗传变化,让自然选择发挥作用。细菌的SOS响应就很好地说明了这一点,因为它可以根据不同环境,发挥这两种功能中的任何一种功能。
参考资料及说明
封面照片: [Source: © vitstudio; Image 134698571 via Shutterstock]
[1] http://atlasgeneticsoncology.org/Educ/DNAID30001FS.html
[2] R. Devoret (1993) Mécanisme de la mutagenèse SOS, Med/Sci vol.3, n°9, I-VII. (in french)
[3] J. Ducau et al (2000) Mutation Research 460:69-80
[4] J. Molinier et al (2006) Nature 442:1046-1049
[5] B.E. Rodgers & K.M. Holmes (2008) Dose response 6:209-221
[6] http://www.perrin33.com/microbiologie/genetique/conjugaison_3.php
[7] Video of Miroslav Radman: https://www.reseau-canope.fr/corpus/video/la-recombinaison-genetique-129.html
[8] D. Anxolabéhère, D. Nouaud & W.J. Miller (2000) Transposable elements and genetic novelties in eukaryotes. Med/Sci, I No.11, vol. 16, I-IX.
[9] 请在 www.lespiedsdansleplat.me中参阅“进化的惊喜:我们祖先中的病毒?”
[10] http://www.cours-pharmacie.com/biologie-moleculaire/reparation-de-ladn.html or http://gec.sdv.univ-paris-diderot.fr/genetique/chapitre9.html
[11] S. Da Re & M.-C. Ploy (2012) Antibiotics and bacterial SOS response. Med Sci (Paris) 28:179-184 (in french)
[12] J. Blázquez, J. Rodrı́guez-Beltrán & I. Matic (2018) Antibiotic-Induced Genetic Variation: How It Arises and How It Can Be Prevented. Annu. Microbiol Rev. 72:209-30
环境百科全书由环境和能源百科全书协会出版 (www.a3e.fr),该协会与格勒诺布尔阿尔卑斯大学和格勒诺布尔INP有合同关系,并由法国科学院赞助。
引用这篇文章: BREGLIANO Jean-Claude (2024年2月23日), 平衡在稳定性和变异性之间的基因组, 环境百科全书,咨询于 2025年1月21日 [在线ISSN 2555-0950]网址: https://www.encyclopedie-environnement.org/zh/vivant-zh/genome-between-stability-and-variability/.
环境百科全书中的文章是根据知识共享BY-NC-SA许可条款提供的,该许可授权复制的条件是:引用来源,不作商业使用,共享相同的初始条件,并且在每次重复使用或分发时复制知识共享BY-NC-SA许可声明。