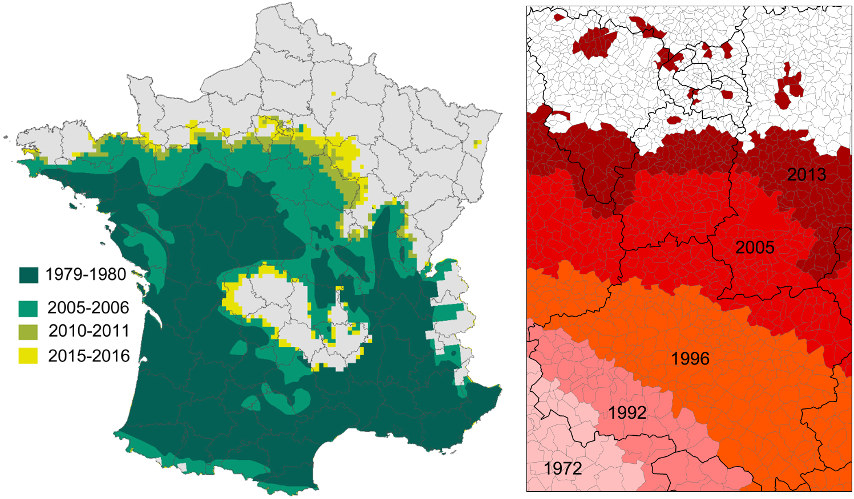
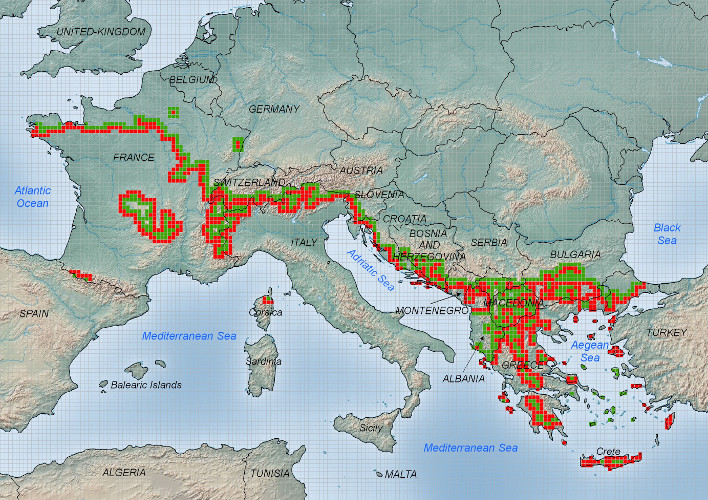
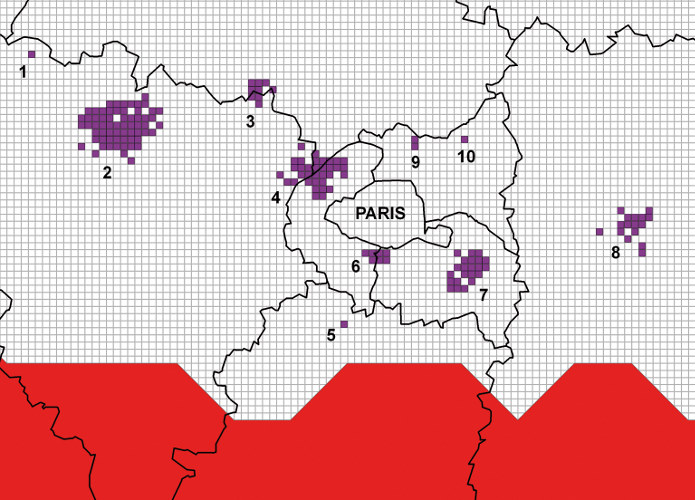
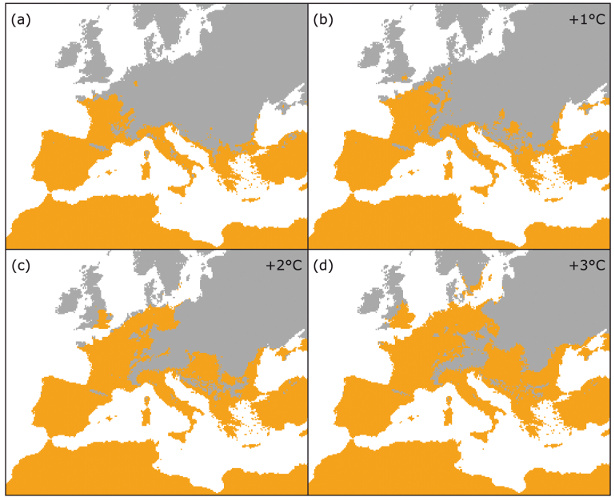
Contrairement à la plupart des autres insectes, le développement larvaire de la processionnaire du pin se déroule durant l’automne et l’hiver et est hautement sensible à de faibles variations de température. Le réchauffement hivernal augmente la survie des chenilles et leur permet de terminer leur développement dans des régions où les conditions climatiques leur étaient préalablement hostiles. Les contraintes climatiques modulant ce développement ont été précisément établies depuis le milieu des années 2000 :
La température létale, en- dessous de laquelle la totalité des colonies de processionnaire ne peuvent survivre, a été estimée à -16°C, valeur qui n’est plus que très rarement atteinte dans la plupart des régions d’Europe de l’Ouest.
Mais au-delà de leur survie, les chenilles doivent aussi s’alimenter pour achever leur développement jusqu’à la nymphose. Elles sortent du nid la nuit pour dévorer les aiguilles de pin, cette sortie nocturne étant soumise à deux conditions [1]:
Le nid doit avoir bénéficié d’une température minimale de 9°C durant la journée (température d’activation)
La température de l’air doit être supérieure à 0°C durant cette nuit (température d’alimentation).
On peut ainsi calculer la mortalité et le nombre de nuits où les chenilles peuvent s’alimenter durant l’hiver, et inversement le nombre de jours consécutifs de famine, qui doit rester limité pour permettre un développement complet [2].
2. A partir de la fin des années 1970, une levée progressive des contraintes thermiques limitant l’aire de répartition de l’insecte
[1] Battisti A., Stastny M., Netherer S., Robinet C., Schopf A., Roques A., et al. (2005) Expansion of geographic range in pine processionary moth caused by increasing winter temperatures. Ecological Applications15, 2084-2096.
[2] Buffo E., Battisti A., Stastny M., Larsson S. (2007). Temperature as a predictor of survival of the pine processionary moth in the Italian Alps. Agricultural and Forest Entomology9, 65–72.
[3] Robinet C., Baier P., Pennerstorfer J., Schopf A., Roques A. (2007). Modelling the effects of climate change on the potential feeding activity of Thaumetopoea pityocampa (Den. & Schiff.) (Lep., Notodontidae) in France. Global Ecology and Biogeography16, 46-471.
[4] Roques A., Rousselet J., Avci M., Avtzis D. N., Basso A. et al. (2015) Climate warming and past and present distribution of the processionary moths (Thaumetopoea spp.) in Europe, Asia Minor and North Africa. pp 81-162 In Roques A. (Ed.) Processionary Moths and Climate Change: An Update. Springer/ Quae.
[5] Battisti A., Avci M., Avtzis D.N., Ben Jamaa M.L., Beradi L. (2015). Natural History of the Processionary Moths (Thaumetopoea spp.): New Insights in Relation to Climate Change. pp. 15-79 In Roques A. (Ed.) Processionary Moths and Climate Change: An Update. Springer/ Quae.
[6] Robinet C, Imbert C.E., Rousselet J., Sauvard D., Garcia J., et al. (2012). Warming up combined with the trade of large trees allowed long-distance jumps of pine processionary moth in Europe. Biological Invasions14, 1557–1569.
[7] Moneo I., Battisti A., Dufour B., Garcia-Ortiz J.C., González-Muñoz M. et al. (2015). Medical and veterinary impact of the urticating processionary larvae. pp. 359- 410 In Roques A. (Ed.) Processionary Moths and Climate Change: An Update. Springer/ Quae.