



Résistance des plantes aux températures extrêmes
PDFLes plantes ont une mobilité réduite et n’ont pas d’autre choix que de faire face aux variations des conditions physiques de leur milieu. Pour s’ajuster et s’adapter à de nouvelles conditions elles doivent percevoir les changements et les traduire en signaux capables de se diriger vers une cible qui réalisera les opérations permettant les changements appropriés au maintien de leur activité. La réponse des plantes à leur environnement implique donc des récepteurs et des molécules de signalisation.
- Les cibles peuvent être (1) le génome nucléaire, chloroplastique ou mitochondrial, (2) des facteurs de transcription, ou encore (3) des activateurs de facteurs de transcription. Des synthèses sont alors initiées ou modulées, voire arrêtées. La réponse touche souvent à la fois le métabolisme et la morphogenèse.
- Noter qu’un signal issu d’une modification perçue seulement en un point de la plante (sur une seule de ses feuilles, par exemple) peut être transmis à la plante entière, et provoquer une réponse globale adaptative qu’on appelle réponse systémique.
Il est difficile de mettre en évidence un récepteur thermique qui, seul, ferait la liaison entre la plante et son environnement : la plante en possède probablement plusieurs en des points clef du métabolisme. Cependant il parait nécessaire que les signaux qu’ils produisent interagissent pour produire la réponse globale appropriée. Noter enfin, que la diversité des systèmes « percevant » les changements de température peut participer à la diversité du monde végétal.
Des recherches utilisant des possibilités de la génétique moléculaire se sont développées dans les deux dernières décades. Elles ont considérablement approfondi nos connaissances sur la réponse aux températures extrêmes. Mais de nombreux points restent à préciser. On présente dans ce focus quelques pistes qui donneront au lecteur un aperçu de la complexité des problèmes posés.
1. Endurcissement et lumière : le thermomètre « phytochromes »
Les interactions entre la lumière et l’endurcissement au froid ont été soulignées depuis longtemps (Lire Effets de la température sur la photosynthèse). Dans certains cas l’endurcissement au froid peut même être obtenu à température ordinaire en modulant la longueur et composition spectrale de la photopériode. Le froid restant cependant nécessaire pour obtenir un endurcissement complet.
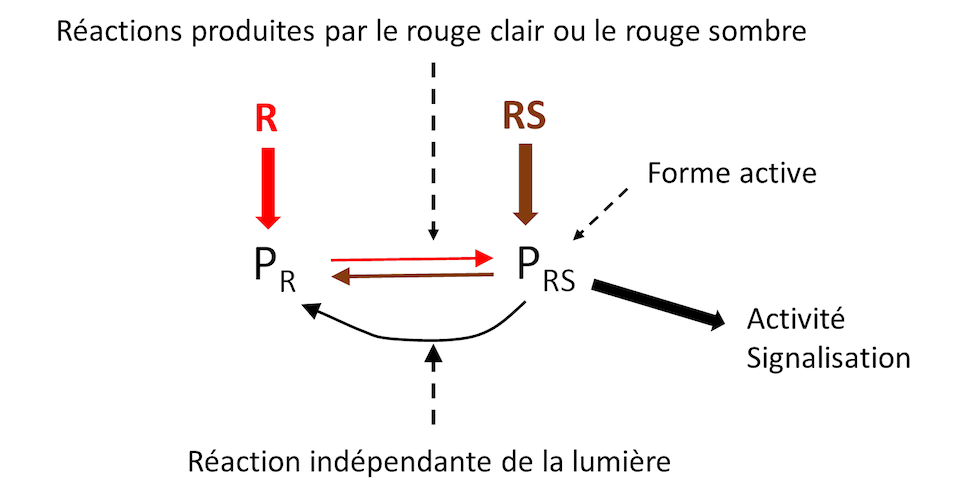
Les phytochromes (P) constituent une famille de chromoprotéines qui permettent à la plante de percevoir la longueur de la photopériode et la composition spectrale dans le rouge (Figure 1). Ils sont bien connus pour être impliqués dans les processus de morphogenèse et de floraison.
- Leur forme inactive (PR) absorbe préférentiellement le rouge clair (R) pour donner une forme active PRS absorbant préférentiellement le rouge sombre (RS).
- Cette réaction est réversible: PRS absorbant du RS donne à nouveau la forme inactive (PR). Le rapport PRS/PR dans la plante exerce un contrôle sur de nombreuses réponses de la plante à la lumière (morphogenèse, floraison etc.) en contrôlant des voies de signalisation aboutissant à une modulation de l’activité de gènes.
- Mais le passage PRS (forme active) vers PR (forme inactive) se fait aussi indépendamment de la lumière. Cette réaction est fortement influencée par la température. Elle peut donc moduler le rapport PRS/PR, et par conséquent l’activité des phytochromes, indépendamment des caractéristiques de l’environnement lumineux.
- Elle constitue un thermomètre capable d’apprécier une différence de 1°C, au moins. [1]
2. La fluidité membranaire : sa relation avec les protéines qui lui sont liées
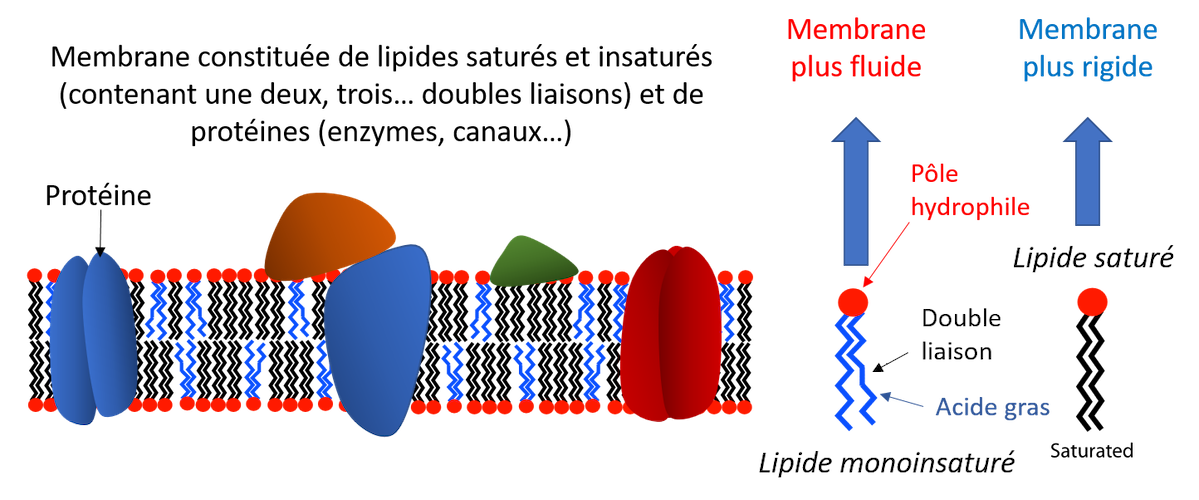
Les membranes biologiques, des bicouches lipidiques, sont composées d’un mélange d’acides gras insaturés (avec une ou plusieurs doubles liaisons), fluides, et d’acides gras saturés (ne possèdent pas de double liaisons), comparativement plus rigides.
Leur fluidité augmente avec la température modifiant non seulement leur perméabilité mais encore l’activité des protéines qui y sont incluses ou qui sont en interaction avec elles.
Cette modification d’activité peut être ainsi à l’origine, par cascades successives, d’une modulation de l’activité de gènes.
Cependant, la fluidité tend à être maintenue, malgré les variations de températures, par une modulation rapide du rapport entre acides gras saturés et non saturés [3], avec une influence sur l’activité des protéines qui y sont incluses.
3. La synthèse d’isoprène
L’isoprène est émis dans l’atmosphère par de nombreuses espèces ligneuses (comme les chênes, les eucalyptus, les conifères…) et herbacées. Plusieurs espèces de mousses (du genre Sphagnum par exemple) et de fougères produisent aussi de l’isoprène. Cependant, de nombreuses plantes n’en n’émettent pas : c’est le cas, par exemple, du Haricot, bien que d’autres papilionacées en produisent.
L’isoprène protège l’appareil photosynthétique [4] contre les chocs thermiques. Cette protection est aussi effective chez les plantes n’émettant pas ou peu d’isoprène, placées dans une atmosphère qui en contient des teneurs compatibles à celles qui sont mesurées à proximité des feuilles de plantes émettrices.
L’isoprène contribuerait à stabiliser rapidement les membranes, mais aussi à réguler la quantité d’espèces actives d’O2. Il agit enfin comme une molécule de signalisation en agissant sur l’ensemble des systèmes impliqués dans la croissance et dans les réponses aux contraintes [5].
4. La production d’espèces actives de l’oxygène (ROS)
La production d’espèces actives d’oxygène est courante chez les plantes soumises à des contraintes de toutes sortes (physiques ou biologiques). Elle se fait, à basses températures comme à températures élevées, principalement dans les chloroplastes, les mitochondries et les peroxysomes.
Par exemple, des espèces actives de l’oxygène (ROS) [6] sont produites en continu lors de la photosynthèse. En effet comme l’absorption de la lumière par les antennes collectrices et son transfert vers les centres réactionnels des photosystèmes sont insensibles à la température, toute baisse de l’assimilation du CO2 produite par les températures extrêmes (basses et hautes) va rendre excédentaire l’énergie lumineuse au niveau des photosystèmes (Lire : Comment les plantes supportent les stress alpins ?)
- Une partie de cette énergie est évacuée sous forme de chaleur au niveau des antennes ;
- Une autre contribue à former des ROS, qui peuvent endommager l’appareil photosynthétique ou déclencher des réponses d’acclimatation.
Les ROS se forment à d’autres étapes du fonctionnement du système de transduction de l’énergie lumineuse en énergie chimique. Elles déclencheraient les réponses adaptatives en interagissant avec les réseaux métaboliques et de signalisation déjà à l’œuvre.
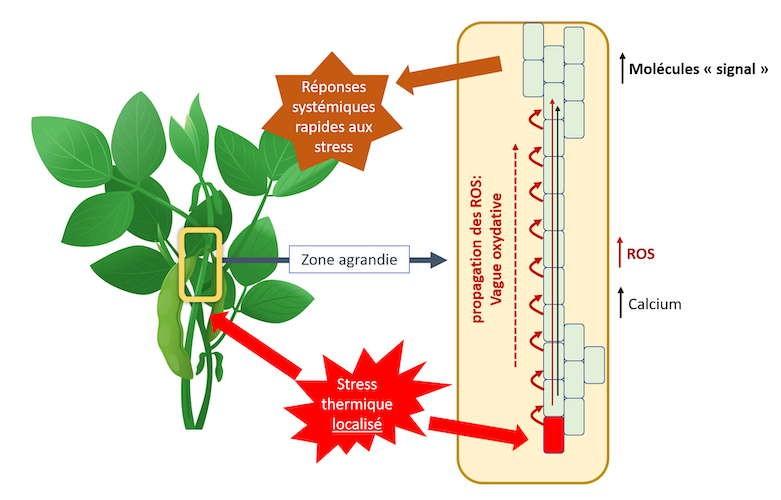
Elles participent aussi, probablement avec le calcium, aux réponses systémiques. On a en effet montré qu’une vague oxydative naissait au point de la plante soumis à une augmentation de température et se propage à la plante entière. Elle a pour origine la production de ROS au point touché par le stress. A partir de ce point se propage une vague oxydative (Figure 3). Cette propagation se fait de cellule en cellule de deux façons :
- Par une voie symplasmique, c’est-à-dire de cytoplasme en cytoplasme via les plasmodesmes et des canaux, les aquaporines par exemple ;
- Par une voie apoplastique, située entre la paroi squelettique et le plasmalemme. Cette dernière voie implique une NADPH oxydase située dans le plasmalemme dont l’activation nécessite du Ca++. Une « vague calcique» accompagne la vague oxydative.
Notes et références
Image de couverture. [Source : © G.Cornic]
[1] Jung, J-H., Domijan, M., Klose, C., Biswas, S.,Ezer, D.,Gao, M., Khattak, A.K., Box, M.S.,Charoensawan, V., Cortijo, S., Kumar, M., Grant, A., Locke, J.C.W., Schäfer, E. , Jaeger, K.E., Wigge, P.A. (2016) Phytochromes function as thermosensors in Arabidopsis. Science 354, 886-889
[2] Catalá, R., Medina, J., Salinas, J. (2011) Integration of low temperature and light signaling during cold acclimation response in Arabidopsis. Proceedings of the National Academy of Sciences 108, 39, 16475-16480
[3] Sung DY, Kaplan F, Lee KJ, Guy CL (2003) Acquired tolerance to temperature extremes. Trends in Plant Science 8, 179-187
[4] Sharkey TD, Chen X, Yeh S (2001) Isoprene increases thermotolerance of fosmidomycine-fed leaves. Plant Physiology 125, 2001-2006
[5] Zuo Z, Weraduwage SM, Lantz AT, Sanchez LM, Weise SE, Wang J, Childs KL, Sharkey TD (2019) Isoprene Acts as a Signaling Molecule in Gene Networks Important for Stress Responses and Plant Growth. Plant Physiology 180, 124-152
[6] Sigle anglais, utilisé communément, pour désigner les espèces réactives de l’oxygène : Reactive Oxygen Species = ROS. Les ROS sont des radicaux qui possèdent un électron célibataire sur leur couche périphérique, caractéristique qui leur permet d’interagir aisément sur leur environnement.
[7] Suzuki N, Devireddy AR, Inupakutika MA, Baxter A, Mille G, Song L, Shulaev E, Rajeev K. Azad, Shulaev V, Mittler R (2015) Ultra-fast alterations in mRNA levels uncover multiple players in light stress acclimation in plants. The Plant Journal 84, 760–772